2008 NERVOV SYSTM Nervov systm Mozek lovka je





 -")











 sensorický neuron obratlovců. Krátké a početné dendrity komunikují se sensory.")





































 Sodíkové kanály jsou zavřené, draselné zůstávají otevřené, aby se mohla")

 dvě brány activation")


























 průnik")









 je místo, kde z těla neuronu vychází axon zde")







 – považována za hlavní neurotransmitter IPSP, zřejmě")








 je")








 – vystýlá lebeční a")








 tvorba energie, vzrušení, příprava těla")







 z metencephalonu (zadního mozku)")
 homeostáza, koordinace pohybů, vedení informací vyšším centrům medulla a pons:")
 všechny sensorické informace, jdoucí do vyššího mozku a všechny motorické")



 koordinuje pohyb, sensorické a")

















")






,")




















- Slides: 192
2008 NERVOVÝ SYSTÉM
Nervový systém Mozek člověka je pravděpodobně nejkomplikovanější struktura na Zemi 1 cm 3 mozkové tkáně obsahuje přes 50 miliónů buněk, navzájem různě propojených v mozku člověka je snad až 1011 buněk spojených 1015 spojů! nervové impulsy běží rychlostí 150 m/s. Endokrinní systém zpravidla reaguje v horizontu minut, hodin nebo dní nervový systém různých skupin živočichů je velmi podobný na úrovni buněk; velmi se ale liší na vyšších úrovních
Přehled nervová soustava: input, integration, output Nervový systém provádí tři hlavní funkce: funkce senzorická - příjem informací z vnějšího i vnitřního prostředí funkce asociační zpracování informací funkce výkonná - pokyny výkonným orgánům (svaly, žlázy)
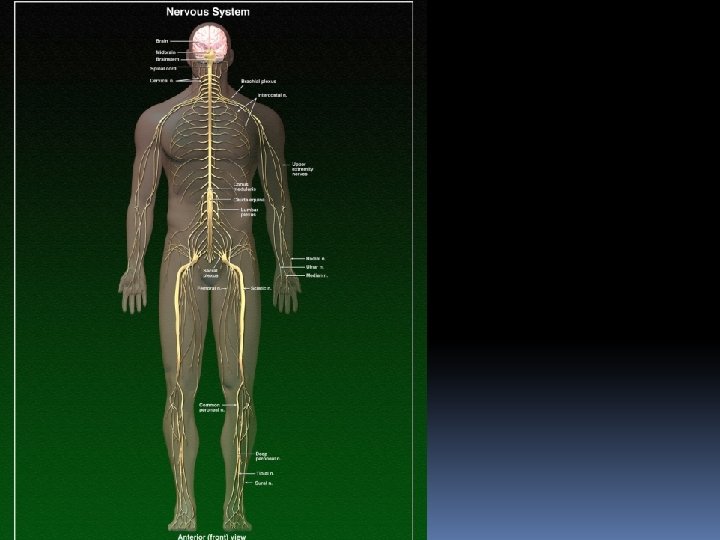
Stavba nervového systému Centrální nervová soustava: mozek + mícha Periferní nervová soustava - zabezpečuje senzorické a motorické signály do CNS a z CNS
Stavba neuronu
Stavba neuronu Tělo - jádro a další organely výběžky dendrity (dendron = strom) - přijímají impulsy a vedou je dovnitř těla buňky axon - obvykle mnohem delší než dendrity. Axon vede impulsy směrem ven z nervové buňky
Axon Vede impulsy ven z buňky neuron může mít mnoho dendritů vždy má ale jen jeden axony, spojující míchu a nohu člověka mohou mít až 1 m délky místo, ve kterém axon nasedá na tělo neuronu se nazývá iniciální segment (axon hillock) opačný konec axonu se nazývá synaptický terminál, ze kterého se při nervovém vzruchu uvolňují chemické látky neurotransmitery
Axon mnoho axonů je kryto izolační vrstvou, zvanou myelinová pochva místo, kde se dva neurony setkávají se nazývá synapse neuron, ze kterého je vzruch vysílán se nazývá presynaptický neuron, který vzruch přijímá se nazývá postsynaptický neuron
Stavba neuronu
Reflexní oblouk
Reflexní oblouk Nejjednodušší typ reflexního oblouku se skládá pouze ze dvou neuronů sensorický neuron - přijímá informaci ze senzoru (změna osvětlení, tlaku, zvuku) a předává ji motorickému neuronu motorický neuron - vede signál efektorové buňce (svalová buňka nebo žláza), která provede odpověď
Patelární reflex Knee-jerk reflex Reflex je způsoben úhozem na šlachu připojené k musculus quadriceps sensorické receptory ve stehnu zachytí změnu napětí ve svalu sensorický neuron převede informaci do míchy v míše informace přeskočí mezi sensorickým a motorickým neuronem aktuální reflex provádějí pouze dva neurony (sensorický a motorický), ale sensorický neuron rovněž komunikuje interneuronem v míše
Patelární reflex Knee-jerk reflex Interneuron inhibuje určité motorické neurony a zabrání stahu flexoru, který je antagonistou m. quadriceps jiné motorické neurony přenesou signál na m. quadriceps, který se stáhne reflexní oblouk je tak jasná a logická věc, že neurofyziologům činilo obtíže přijmout roli hormonální soustavy, neboť nebylo jasné, proč si nervová soustava nevystačí sama…?
Reflexní oblouk
Reflexní oblouk Většinou je vloženo mezi sensorický a motorický neuron mnoho interneuronů, které mezi sebou komunikují. Interneurony zpracovávají sensorické informace a připravují optimální behaviorální odpověď i u velmi jednoduchých živočichů jsou interneurony neustále aktivní, a „povídají si“ mezi sebou velké množství interneuronů v mozku člověka rozhoduje, které chování nakonec nastane pokud se například vůlí rozhodneme neuskutečnit patelární reflex, interneurony mozku vyšlou signál do míchy a i po klepnutí kladívkem zůstane koleno nehybné
Šedá a bílá hmota Těla interneuronů a motorických neuronů tvoří šedou hmotu centrálního nervového systému v míše je šedá hmota uvnitř a bílá vnější bílá hmota míchy je tvořena sensorickými a motorickými axony těla sensorických neuronů se však nachází vně míchy, kde tvoří útvary zvané zadní kořeny míchy. Ganglion je shluk těl neuronů, často podobné funkce, umístěný na periferii NS podobné útvary v mozku se nazývají jádra (nuclei)
Různé typy neuronů Těla neuronů a dendrity černě, axony fialově. Neuron na obr. b dole má až 100 000 synapsí!
Různé typy neuronů (a) sensorický neuron obratlovců. Krátké a početné dendrity komunikují se sensory. Dlouhý axon vede vzruchy do CNS. Tělo buňky je umístěno na axonovém vlákně. (b) dva typy interneuronů z mozku savce. Na horním obrázku je dobře vidět rozvětvený axon (c) motorický neuron bezobratlých. Tělo neuronu zde nasedá na dendrity
Typy nervových okruhů informace z jednoho zdroje, např. z oka dostává do několika míst mozku, tedy z jednoho presynaptického neuronu jde do několika postsynaptických informace z více zdrojů jdou do jednoho místa v mozku. Např. vidění, hmat a sluch pomáhají identifikovat objekt. Z mnoha presynaptických neuronů jde tedy informace do jednoho postsynaptického v třetím případě může jít informace kruhem. Takto snad může být uchována informace v lidském mozku - krátkodobá paměť
Gliové buňky Je jich 10 - 50 x víc než nervových buněk (Glia = lepidlo) donedávna se mělo za to, že mají pouze pomocnou roli; výzkumy posledních let ukázaly, že existují synapse i mezi gliovými buňkami a neurony v mozku a míše je několik typů gliových buněk v embryu gliové buňky (radial glia) razí cestu, po které porostou neurony z nervové trubice za vzniku CNS
Gliové buňky V dospělém CNS gliové buňky zvané astrocyty poskytují strukturální a metabolickou podporu neuronům astrocyty jsou též odpovědné za tvorbu těsných spojů (tight junctions) mezi kapilárami v mozku. Výsledkem je bariéra mezi mozkem a krví, která zabraňuje proniknutí většiny substancí z krve do mozku. Výsledkem je přesná kontrola extracelulárního prostředí mozkových buněk výzkumy posledních let naznačují, že astrocyty komunikují jeden s druhým a s neurony pomocí chemických signálů
Astrocyty
Gliové buňky oligodendrocyty a Schwanovy buňky Oligodendrocyty jsou v CNS Schwanovy buňky jsou v PNS Obojí jsou gliové buňky, které vytvářejí myelinový obal kolem axonů mnoha neuronů
Gliové buňky oligodendrocyty a Schwanovy buňky Neurony jsou myelinované tehdy, když v jejich vývoji se Schwanova buňka nebo oligodendrocyt rostou kolem axonu a v mnoha vrstvách jej obtáčejí Membrány jsou lipidové, což má za následek velmi slabou vodivost těchto membrán Výsledkem je izolace axonů, funkčně podobné umělohmotné izolaci elektrických drátů
Gliové buňky oligodendrocyty a Schwanovy buňky Při roztroušené skleróze se myelinový obal axonů rozpadá, což má za následek nesprávné vedení elektrických impulsů v axonech Výsledkem je postupná ztráta koordinace Na fotografii jsou vidět světlé plaky způsobující roztroušenou Sklerózu. Obrázky jsou snímány ve dvoutýdenních intervalech
Hematoencefalická bariéra = bariéra mezi krví a CNS „tight junctions“ mezi endotelovými buňkami kapilár Absence fenestrací – na rozdíl od obvyklých tělních kapilár nemají mozkové kapiláry fenestrovanou („proděravělou“) stěnu Omezení pinocytózy - exocytózy a endocytózy Astrocyty vytvářejí kolem kapilár val
Povaha nervových signálů Všechny buňky mají rozdíl v elektrickém náboji na obou stranách plasmatické membrány Tomuto rozdílu se říká membránový potenciál Membránový potenciál lze měřit voltmetrem, jehož jedna elektroda je v buňce a druhá na vnější straně membrány Elektroda mimo buňku nese název referenční elektroda
Membránový potenciál V živočišné buňce se membránový potenciál pohybuje mezi – 50 m. V až – 100 m. V Znaménko „-“, minus, signalizuje, že vnitřek buňky nese záporný náboj Neuron v klidu má membránový potenciál okolo – 70 m. V Tento potenciál je jmenován klidový potenciál
„Giant axons“ Neurony některých bezobratlých, jako např. sépií, jsou neobvykle velké a lze na nich proto snadno studovat vedení nervového vzruchu Některé neurony sépií mají průměr až 1 mm! – lze proto do nich snadno vložit elektrody a provést měření. Řada objevů ze základního výzkumu byla provedena právě na obřích axonech těchto hlavonožců
Jak neuron udržuje membránový potenciál? Membránový potenciál existuje díky nerovnoměrnému rozložení iontů na obou stranách plasmatické membrány Tento rozdíl udržuje membránový přenašeč, zvaný sodíko-draslíková pumpa
Jak neuron udržuje membránový potenciál? Uvnitř buňky je hlavním iontem draslík K+, ačkoli jsou zde přítomny i ionty sodíku Na+ Mimo buňku je situace opačná, hlavním iontem je zde sodík Na+, ačkoli jsou zde i ionty K+ Uvnitř buňky jsou hlavními anionty proteiny, aminokyseliny, sulfáty, fosfáty a další látky, symbolizované na obrázku jako AProtože ionty nesou náboj, fosfolipidová dvojvrstva je pro ně nepropustná
Jak neuron udržuje membránový potenciál? Uvnitř buňky jsou v nízké koncentraci přítomny rovněž chloridové anionty Cl Mimo buňku jsou Cl- hlavními anionty I v klidu existuje stálá difuse K+ směrem ven z buňky a iontů Na+ dovnitř. Velikost šipek symbolizuje velikost difuse. Na+/K+ pumpa vyrovnává gradient v opačném směru.
Na+/K+ pumpa Na/K pumpa přenáší vždy 3 ionty Na+ směrem ven a zároveň 2 ionty K+ směrem dovnitř buňky. Energii dodává ATP.
Iontové kanály Pro průnik membránou musí být ionty být buď přeneseny Na/K pumpou, nebo mohou přejít iontovými kanály, které jsou specifické pro každý druh iontu některé kanály umožňují průchod pouze Na+ iontů, jiné K+ iontů a ještě jiné pouze Cl- iontů podle toho, kolik má plasmalema iontových kanálů jsou určeny její vlastnosti Různé plasmatické membrány se tak mohou chovat zřetelně odlišně
Iontové kanály Většina buněk, včetně neuronů, má mnohem větší propustnost pro K+ než pro Na+ Plasmalema neuronů je pro K+ až 50 x propustnější než pro Na+ což nasvědčuje tomu, že je v membráně mnohem více K+ kanálů než Na+ kanálů protože anionty jsou většinou velké molekuly (proteiny apod. ), které nemohou proniknout ven, vnitřek axonu nese vždy záporný náboj
Elektrický a chemický gradient Ionty se pohybují tak, aby se nastolila rovnováha, tedy z míst o větší koncentraci do míst o menší koncentraci (chemický gradient) a rovněž tak, aby se vyrovnal elektrický náboj (elektrický gradient) Ionty K+ jsou taženy směrem ven z buňky chemickým gradientem, ale naopak směrem dovnitř gradientem chemickým
Elektrický a chemický gradient kdyby K+ byl jediný iont v buňce, chemický i elektrický gradient by se časem vyrovnal na hodnotě – 85 m. V plasmalema je mnohem méně propustná pro Na+ ionty, avšak tyto jsou taženy dovnitř buňky jak chemickým, tak elektrickým gradientem výsledný potenciál je tedy o něco menší, typicky okolo - 70 m. V.
Elektrický a chemický gradient kdyby se difuse obou iontů ponechala svému osudu, časem by Na+ proniklo do buňky a K+ by proniklo ven klidové potenciál proto neustále vyrovnává Na/K pumpa, které čerpá Na+ ven a K+ dovnitř
Iontové kanály Všechny buňky mají membránový potenciál pouze svalové a nervové buňky ale mají schopnost změn tohoto potenciálu změna tohoto potenciálu může vyústit v elektrický impuls některé iontové kanály jsou otevřené neustále jiné jsou uzavřené, a mohou se otevřít nebo zavřít po obdržení stimulu (gated ion channels)
Iontové kanály Stimul může přijít jak z vnějšího prostředí (světlo pro oko nebo chvění vzduchu pro sluch), nebo z prostředí vnitřního (elektrické nebo chemické impulsy pro interneurony) některé iontové kanály se otevírají nebo zavírají na chemický stimul, jako jsou neurotransmittery uvolněné ze synaptického terminálu (chemically-gated ion channels) jiné se otevírají a zavírají v souvislosti se změnou membránového potenciálu (voltage-gated ion channels)
Navíc jeden typ kanálu propouští pouze jeden druh iontu existuje tak sodíkový chemically-gated ion channel a sodíkový voltage-gated ion channel, stejně jako draslíkový chemically-gated ion channel a draslíkový voltage-gated ion channel
Hyperpolarizace a depolarizace Je-li dendrit stimulován neurotransmitterem, záleží na tom, jaký typ iontového kanálu bude otevřen. Může tak dojít k hyperpolarizaci nebo depolarizaci
Hyperpolarizace = zvětšení membránového potenciálu. Jednou z možností je otevření K+ kanálu, čímž se zvýší jeho průnik ven z buňky po chemickém gradientu, a vnitřek buňky se stane negativnějším
Depolarizace = snížení membránového potenciálu. Jednou z možností je otevření Na+ kanálů. Sodík proniká po chemickém i elektrickém gradientu do buňky a membránový potenciál se snižuje
Hyperpolarizace a depolarizace Změny v elektrickém napětí způsobené hyperpolarizací a depolarizací jsou graduované: velikost změny závisí na velikosti stimulu větší stimul otevře více kanálů a způsobí větší změnu v membránovém potenciálu
Akční potenciál Depolarizace membrány je ovšem graduovaná pouze k určitému bodu, zvanému prahový potenciál pokud depolarizace dosáhne tohoto prahového potenciálu, vznikne nový typ odpovědi, zvaný akční potenciál
Akční potenciál může v neuronu vzniknout pouze v axonu. . Může jej spustit depolarizace dendritu nebo těla buňky, odkud se rozšíří až do axonu Prahový potenciál je typicky o 15 až 20 m. V menší než klidový potenciál V axonu se tedy jedná o potenciál v rozmezí -50 m. V až -55 m. V
Hyperpolarizace, depolarizace, akční potenciál
Akční potenciál je negraduovaný; jedná se o odpověď typu vše-nebo-nic velikost akčního potenciálu je tedy nezávislá na velikosti depolarizace, která jej vyvolala. Jakmile je jednou akční potenciál spuštěn, membránový potenciál prochází stereotypní sekvencí změn
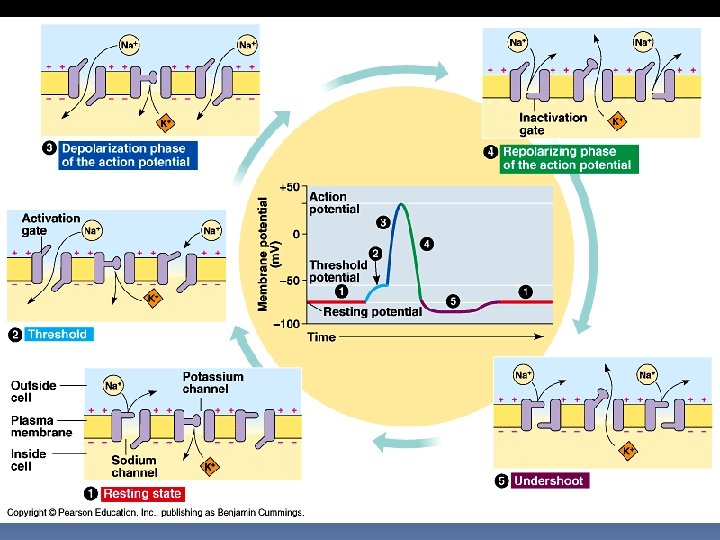
Akční potenciál
Akční potenciál Při akčním potenciálu dojde k rychlému přepólování membrány, s kladným nábojem uvnitř a záporným nábojem vně buňky tento převrat je následován repolarizací, kdy se náboje vrátí na svou původní hodnotu celá událost netrvá déle než milisekundy akční potenciál je příkladem pozitivní zpětné vazby - depolarizace vedoucí k prahovému potenciálu spustí ještě mohutnější depolarizaci akční potenciál
Akční potenciál Klidový stav. Sodíkové i draslíkové kanály jsou uzavřeny, a v axonu je udržován klidový potenciál
Akční potenciál Stimul otevírá Na+ kanály. Je-li dosaženo prahového potenciálu, dochází k otevření ještě dalších sodíkových kanálů a vzniká akční potenciál
Akční potenciál Depolarizační fáze. Sodíkové ionty ve velkém množství pronikají rychle dovnitř. Draslíkové kanály jsou uzavřeny. Výsledkem je kladný náboj uvnitř a záporný vně.
Akční potenciál Repolarizační fáze. Sodíkové kanály se uzavírají a draslíkové se otevírají. K+ ionty opouští buňku, čímž se vnitřek buňky stává zápornější než vnějšek
Akční potenciál Undershoot (hyperpolarizace) Sodíkové kanály jsou zavřené, draselné zůstávají otevřené, aby se mohla dokončit repolarizace mebrány. Po 1 - 2 milisekundách je klidový potenciál obnoven a axon je připraven odpovědět na další stimul
Akční potenciál Vznik akčního potenciálu je umožněn existencí voltage-gate ion channels, které se uzavírají a otevírají v souvislosti se změnou membránového potenciálu draslíkový voltage-gated ion channel je uzavřen v klidovém potenciálu a otevírá se pomalu v odpověď na depolarizaci
Akční potenciál sodíkový voltage-gated ion channel má (na rozdíl od draselného) dvě brány activation gate, která je za klidu uzavřená a otevírá se rychle v závislosti na depolarizaci inactivation gate, která je v klidu otevřena a zavírá se pomalu v odpověď na depolarizaci
Akční potenciál Po dosažení akčního prahového potenciálu: K+ brána se pomalu otevírá a K+ ionty pomalu opouští buňku aktivační brána Na+ kanálů je hned a široce otevřena a Na+ ionty prudce vtrhávají do buňky inaktivační brána Na+ kanálů se začíná pomalu uzavírat
Akční potenciál Rychlá depolarizace je následována repolarizací, pík akčního potenciálu je vysoký a ostrý. Repolarizace je způsobena: sodíkové kanály se uzavírají draselné jsou otevřeny, čímž K+ se dostává ven a vnitřek buňky se stává zápornějším díky proudění K+ ven z buňky nastává na okamžik „přestřelení“ membránového potenciálu, dochází na chviličku k hyperpolarizaci membrány
Akční potenciál Během této hyperpolarizace jsou obě brány sodíkových kanálů zavřeny pokud během této fáze neuron obdrží další depolarizační stimul, není schopen na něj zareagovat, neboť inaktivační brána se ještě neměla čas otevřít toto období, ve kterém je neuron necitlivý k depolarizaci , se nazývá refraktorní perioda
Akční potenciál Neuron tedy neumí dopovídat na silné a slabé stimuly: akční potenciál je vždy týž pokud je ovšem stimul velmi silný, projeví se sérií akčních potenciálů, jdoucích tak rychle za sebou, jak to refraktorní perioda dovolí.
Nervový impuls postupuje axonem Akční potenciál ve skutečnosti nepostupuje, ale opakovaně se regeneruje v průběhu axonového vlákna regenerace akčního potenciálu je způsobena tím, že vpád Na+ iontů do buňky způsobí elektrický proud, který depolarizuje i sousední oblasti, a to tak silně, že dojde i zde ke vzniku prahového potenciálu, čímž se spustí nový akční potenciál
Nervový impuls postupuje axonem Akční potenciál lze přirovnat k shození první kostky domina. Hyperpolarizace refraktorní periody zabraňuje tomu, aby se akční potenciál šířil směrem vzad. Axon tedy lze přirovnat k ulici, po které jezdí auta jen jedním směrem
Změna membránového potenciálu může vyústit v elektrický impuls
Rychlost propagace akčního potenciálu - bezobratlí Závisí na průměru vlákna: čím je vlákno větší, tím vzrůstá i rychlost vedení vzruchu. U silného axonu může depolarizace dosáhnout dále než u tenkého. Tenké axony šíří vzruch rychlostí několika centimetrů za vteřinu. Giant axons sépií a humrů až 100 m/s tyto obří axony odpovídají za rychlý pohyb ocasu směrem zad při útěkové reakci humrů a raků
Rychlost propagace akčního potenciálu - obratlovci Mnoho axonů obratlovců je myelinováno a to buď oligodendrocyty (CNS), nebo Schwanovými buňkami (PNS) voltage-gated ion channels jsou koncentrovány především v Ranvierových zářezech, což jsou jediné části axonu komunikující s intesticiální tekutinou proudění iontů dovnitř a ven se tedy realizuje pouze v těchto Ranvierových zářezech
Rychlost propagace akčního potenciálu - obratlovci Akční potenciál se tedy nepropaguje postupně, ale spíše skokem od jednoho Ranvierova zářezu ke druhému, kde stimuluje depolarizaci a nový akční potenciál tento mechanismus je zván saltatorní kondukce (vedení) vzruchu v myelinovaných neuronech se může saltatorní kondukcí vzruch šířit rychlostí až 150 m/s
Rychlost propagace akčního potenciálu - obratlovci Proud iontů se pohybuje v akčním potenciálu vnitřkem axonu (modré šipky) až k dalšímu Ranvierovu zářezu, kde způsobí nový akční potenciál. Akční potenciál tak „skáče“ od jednoho Ranvierova zářezu ke druhému.
Chemická nebo elektrická komunikace mezi buňkami se uskutečňuje díky synapsím Synapse jsou zvláštním typem spojů mezi buňkami, kontrolují komunikaci mezi neuronem a jinou buňkou synapse se nacházejí mezi: neuronem a neuronem sensorem a sensorickým neuronem motorickým neuronem a svalovou buňkou neuronem a buňkou dané žlázy
Chemická nebo elektrická komunikace mezi buňkami se uskutečňuje díky synapsím Synapse jsou dvojího typu: elektrické synapse chemické synpse
Elektrické synapse Elektrická synapse dovoluje přechod akčního potenciálu přímo z buňky presynaptické na buňku postsynaptickou buňky jsou spojeny spoji zvanými „gap junctions“ mezibuněčné kanály tak dovolují tok iontů mezi oběma neurony obří axony humrů a dalších korýšů jsou spojeny elektrickými synapsemi, což umožňuje propagaci vzruchu bez ztrát na rychlosti a intenzitě
Některé typy spojů mezi buňkami opakování
Elektrické synapse v CNS obratlovců synchronizují aktivitu neuronů odpovědných za rychlé stereotypní jednání… … například rybám v rychlém pohybu ocasem při „útěku“ před predátorem podobně u raků při prudkém pohybu vzad
Chemické synapse Jsou mnohem častější než elektrické synapse úzký prostor, zvaný synaptická štěrbina, odděluje presynaptickou buňku od postsynaptické díky této štěrbině buňky nejsou propojeny, a akční potenciál nemůže přeskočit z jedné buňky na druhou namísto toho nastává série událostí, ve kterých je akční potenciál proměněn na chemický signál, který překoná synaptickou štěrbinu a v postsynaptické buňce je konvertován zpět na akční potenciál
Chemická synapse
Synaptický terminál
Chemické synapse Cytoplasma synaptického terminálu obsahuje řadu váčků zvaných synaptické vesikly každý váček obsahuje tisíce molekul neurotransmitterů, chemické látky, která bude uvolněna do synaptické štěrbiny ačkoli je známo mnoho druhů neurotransmitterů, většina neuronů vylučuje vždy jen jeden typ neurotransmitteru
Chemické synapse postsynaptický neuron však může přijímat signály od mnoha presynaptických neuronů, které vylučují ze synaptických terminálů mnoho druhů neurotransmitterů když akční potenciál doputuje na konec axonu, způsobí depolarizaci presynaptické membrány, což je plasmalema synaptického terminálu obrácená k synaptické štěrbině podstatnou roli zde hrají ionty Ca++
Chemické synapse Depolarizace presynaptické membrány způsobí rychlý vpád Ca++ iontů do cytoplasmy synaptického terminálu díky vápníkovým voltagegated ion channels náhlý a prudký vzestup koncentrace Ca++ iontů v synaptickém terminálu způsobí, že vesikly splynou se synaptickou membránou a exocytózou vyloučí neurotransmittery do synaptické štěrbiny na jediný akční potenciál tak reagují zároveň unisono stovky váčků
Chemické synapse Neurotransmittery překonají difúzí krátkou vzdálenost k postsynaptické membráně, což je plasmalema dendritu nebo těla postsynaptické buňky tato plasmalema má v sobě zabudované receptory pro neurotransmittery tyto receptory jsou součástí selektivních iontových kanálů, které se mohou otevírat a zavírat a regulovat tak průnik iontů do buňky
Chemické synapse Pokud se neurotransmitter naváže na receptor, způsobí se otevření iontového kanálu buď pro K+ nebo pro Na+ nebo pro Cl iontové kanály postsynaptické membrány jsou tedy chemical-gated ion channels, na rozdíl od voltage-gated ion channels axonového vlákna
Chemické synapse Průnik iontů těmito kanály změní membránový potenciál postsynaptické buňky podle typu receptoru a podle druhu přenášeného iontu je způsobena buď hyperpolarizace nebo depolarizace neurotransmitter je však velmi rychle odbourán, buď enzymaticky, nebo je vstřebán sousedními buňkami tím je zaručeno, že efekt neurotransmitterů na postsynaptickou buňku bude rychlý a precizní
Chemické synapse Například neurotransmitter acetylcholin je rychle degradován enzymem cholinesterázou, který je přítomen jak v synaptické štěrbině, tak i v postsynaptické membráně impuls může být veden pouze jedním směrem: pouze v synaptickém terminálu jsou vesikly s neurotransmittery, a pouze v postsynaptické membráně jsou receptory pro tyto neurotransmittery
Chemické synapse Když akční potenciál depolarizuje membránu synaptického terminálu, tím se spustí (1) průnik Ca++ do buňky, což způsobí (2) splynutí synaptických vesiklů s membránou presynaptického neuronu. Vesikly (3) uvolní neurotransmitter do synaptické štěrbiny, které se navážou na receptory v postsynaptické membráně (4). Molekuly neurotransmitterů jsou rychle degradovány (5) buď enzymy, nebo jsou pohlceny sousedními neurony
Neurální integrace se objevuje již na buněčné úrovni Jediný neuron může obdržet informaci od sousedních neuronů díky tisícům synapsí, z nichž některé z nich mohou neuron excitovat, jiné inhibovat například u patelárního reflexu jsou synapse mezi sensorickými neurony a motorickými neurony, stimulujícícmi m. quadriceps ke stahu excitační a naopak synapse mezi interneurony a motorickými neurony inervujícími flexor inhibiční
Integrace mnoha synaptických signálů
EPSP excitatory postsynaptic potential Excitační synapse otevře kanály tak, že Na+ proniká do buňky (po obou gradientech) a K+ buňku opouští (po chemickém gradientu). Tím vzrůstá kladný náboj vnitřku buňky, membrána se depolarizuje a blíží se k prahovému potenciálu v tomto případě je elektrická změna, způsobnená navázáním neurotransmitteru na receptor zvaná excitační postsynaptický potenciál, EPSP
IPSP inhibitory postsynaptic potential Inhibiční synapse otevírají kanály pro K+, pro Clnebo pro oba tyto ionty. K+ buňku opouští a chloridové buňky naopak proudí dovnitř buňky, oba po chemickém gradientu. Tím dochází k hyperpolarizaci membrány a snižuje se pravděpodobnost vzniku akčního potenciálu. Tato elektrická změna je zvány inhibiční postsynaptický potenciál, IPSP
EPSP a IPSP EPSP i IPSP jsou graduované potenciály, které se liší svou intenzitou, počtem neurotransmitterů navázaných na receptory postsynaptické membrány lokální elektrická změna membrány, depolarizace nebo hyperpolarizace, trvá pouze několik milisekund, díky tomu, že neurotransmittery jsou rychle odbourány nebo vstřebány
Sumace
EPSP a IPSP aby se postsynaptická buňka „zažehla“, musí být EPSP natolik silné, aby se u počátku axonu dosáhla prahová hodnota - 50 m. V jediný EPSP zpravidla není natolik silný, aby způsobil akční potenciál postsynaptické buňky. Pokud je ovšem EPSP více, nebo pokud menší množství synaptických terminálů uvolňuje opakovaně neurotransmittery, může se dosáhnout prahového potenciálu tento aditivní efekt EPSP je zván sumace
Sumace Existují dva typy sumace: časová prostorová při časové sumaci jdou EPSP tak rychle po sobě, že postsynaptická membrána nemá čas se vrátit ke klidovému stavu při prostorové sumaci působí najednou řada synaptických terminálů, náležejících zpravidla různým neuronům
Sumace rovněž nastává u IPSP, kdy může dojít k hyperpolarizaci postsynaptické membrány tak velkému, že by jedna IPSP jej nemohla způsobit EPSP a IPSP tak působí antagonisticky proti sobě
Sumace Iniciální segment (axon hillock) je místo, kde z těla neuronu vychází axon zde se sumují všechny EPSP a IPSP, které buňka v daném okamžiku obdrží pokud účinek všech EPSP převáží hyperpolarizační účinek IPSP, buňka se „zažehne“ a vyšle svůj vlastní akční potenciál o několik milisekund později, pokud trvá stav depolarizace, může buňka po refraktorní periodě vyslat další akční potenciál
Stejný neurotransmitter může mít odlišný účinek na různé neurony některé neurotransmittery pracují rychle a přesně, pouze v průběhu milisekund účinek jiných trvá delší dobu, až několik minut v některých případech může účinek trvat ještě déle (ovlivnění nálady, soustředěnosti, vzrušení)
Nejčastější neurotransmittery
Acetylcholin je jeden z nejčastějších neurotransmitterů bezobratlých i obratlovců v CNS obratlovců může působit excitačně i inhibičně působí excitačně mezi motorickým neuronem a svalem Na tytéž svalové receptory se navazuje nikotin naopak v srdečním svalu je acetylcholin uvolňován parasympatikem, otevírá K+ kanály a inhibuje enzym adenylcyklázu, čímž zpomaluje sílu a frekvenci srdečních svalů
Acetylcholin Na fotografii ze světelného mikroskopu jsou vidět neurosvalové ploténky. Hlavním neurotransmitterem je zde acetylcholin
Biogenní aminy: Noradrenalin, dopamin, serotonin jsou odvozeny od aminokyselin katecholaminy jsou odvozeny od aminokyseliny tyrozinu; patří sem adrenalin a noradrenalin. Oba působí též jako hormony. Chemicky velmi podobný je i dopamin Dopamin a serotonin jsou uvolňovány na mnoha místech mozku a ovlivňují náladu, spánek, schopnost učení deprese mohou být úspěšně léčeny zvýšenými dávkami noradrenalinu a serotoninu nedostatek dopaminu je spojován s Parkinsonovou chorobou, jeho přebytek se schizofrenií
Biogenní aminy: Noradrenalin, dopamin, serotonin je odvozen od aminokyseliny tryptofanu tyto tzv. biogenní aminy často ovlivňují i enzymatickou aktivitu v postsynaptických buňkách LSD a meskalin způsobují halucinogenní efekty tím, že se navazují na serotoninové a dopaminové receptory v mozku
Účinky drog na bezobratlé normální síť pavouka LSD pavouk po aplikaci marihuany Prášky na spaní
Další neurotrasmittery kyselina γ- amino máselná (GABA) – považována za hlavní neurotransmitter IPSP, zřejmě tím, že otevírá kanály pro ionty Cl glycin, kyselina asparagová, kyselina glutamová substance P – považována za hlavní látku zprostředkovávající pocity bolesti endorfiny – tlumí pocity bolesti během fyzického či emočního stresu, porodu atd. Navíc způsobují pocity euforie, ovlivňují produkci moči a hloubku dýchání. Na jejich receptory v CNS se váže heroin a morfin.
Neurotransmittery na bázi plynů: NO a CO některé neurony v PNS i CNS užívají jako neurotransmittery NO a CO během sexuálního vzrušení u mužů vylučují určité neurony oxid dusnatý v erektilních tkáních penisu. Díky tomu se buňky hladkého svalstva v stěnách kapilár erektilní tkáně roztahují a houbovitá erektilní tkáň se plní krví, čímž je způsobena erekce. Léčivo Viagra inhibuje enzym, který snižuje aktivitu NO, čímž udržuje erekci.
Neurotransmittery na bázi plynů: NO a CO mnoho buněk vylučuje molekuly plynů v odpověď na chemické signály například acetylcholin, produkován neurony ve stěnách krevních kapilár stimuluje endotelové buňky těchto cév k tvorbě NO. NO pak způsobuje uvolnění těchto buněk, což způsobuje roztažení kapilár
Neurotransmittery na bázi plynů: NO a CO Objev tohoto mechanismu konce 80. let XX. stol. vysvětlil již přes sto let známý účinek nitroglycerinu na léčbu anginy pectoris (bolest srdce spojená s nedostatkem krve v srdečním svalu). Enzymy mění nitroglycerin na NO, čímž se roztahují a uvolňují kapiláry, zásobující srdce krví.
Neurotransmittery na bázi plynů: NO a CO na rozdíl od ostatních neurotransmitterů nejsou plyny uschovávané ve vesiklech, ale jsou tvořeny pouze „na požádání“ difúzí se dostávají k nejbližším cílovým buňkám, kde způsobí změnu, a jsou v zápětí odbourány, to vše během sekund NO pracuje tak jako mnoho hormonů: sám do buňky nevstupuje, ale způsobí změnu v membránovém receptoru, čímž podnítí syntézu druhého přenašeče (second messenger)
Neurotransmittery na bázi plynů: NO a CO oxid uhelnatý je syntetizován enzymem oxigenázou, jak v CNS tak i v PNS v mozku CO reguluje uvolňování hormonů z hypotalamu v PNS působí CO jako neurotransmitter – a to inhibičně; hyperpolarizuje stěny buněk hladkých svalů ve stěně střeva
Evoluce a diversita nervové soustavy
Schopnost buněk reagovat na prostředí se objevila před miliardami let prokaryota se umí pohybovat chemotakticky k potravnímu zdroji nebo prchat z nepříznivého prostředí od doby kambrické exploze (cca 530 miliónů let) se živočichové umí pohybovat relativně rychle a odpovídat na environmentální stimuly
Nervové systémy mají různý stupeň organizace v celé říši Animalia se akční potenciál pohybuje stejným způsobem. Rozdíly jsou v organizaci nervových buněk rozdíly tedy nejsou v základních stavebních kamenech, neuronech, ale v tom, jakým způsobem jsou sestaveny
Nervové systémy mají různý stupeň organizace houbovci nervovou soustavu nemají u nezmara (Cnidaria) je difúzní nervová soustava, kontrolující kontrakce a uvolnění láčky cephalizace, seskupení neuronů v přední části těla, umožňuje komplexnější chování; ploštěnka, která má nervové provazce a malý mozek, je tak již schopna složitějšího pohybu; je tak nejjednodušším tvorem s CNS
Nervové systémy mají různý stupeň organizace u komplexnějších bezobratlých, jako jsou Annelida a Insecta je chování kontrolováno složitějším mozkem a břišními nervovými páskami s pravidelně uspořádanými ganglii u obratlovců běží mícha spíš dorzálně, a chybí zde ganglia
Nervové systémy mají různý stupeň organizace Mollusca jsou příkladem toho, jak v rámci jedné skupiny organizace nervového systému souvisí se způsobem života. chroustnatka (Polyplacophora) má při svém přisedlém způsobu života cephalizaci jen naznačenou a velmi sporé sensorické orgány naopak krakatice, sépie a chobotnice mají nervovou soustavu srovnatelnou s obratlovci velké oči spolu s mohutným mozkem, a obřími axony odpovídá pohyblivému způsobu života těchto dravců
Evoluce a diversita nervové soustavy
Mozek člověka
Mozek člověka Snímek byl pořízen technikou magnetické rezonance. Barevně jsou vyznačeny oblasti mozku, které jsou aktivní, když pokusná osoba mluví
Nervový systém obratlovců rozdělení na CNS a PNS vývoj z duté nervové trubice vysvětluje fakt, že mícha i mozek obsahují prostory naplněné mozkomíšním mokem CNS – mozek a mícha, vznikly embryonálně z duté nervové trubice, které je charakteristickým znakem všech Chordat. Mozkové nervy, míšní nervy a ganglia mimo CNS tvoří dohromady PNS
Nervový systém obratlovců tzv. centrální kanál míchy je napojen na čtyři mozkové komory, vyplněné mozkomíšním mokem mozkomíšní mok vzniká v mozku jako filtrát z krve mozkomíšní mok obsahuje živiny, hormony, bílé krvinky mozkomíšní mok má rovněž mechanickou funkci, tlumí nárazy a působí tak jako jakýsi polštář
Komory, šedá hmota a bílá hmota Čtyři mozkové komory, znázorněné modře, jsou vyplněny mozkomíšním mokem
Obaly CNS chrání a vyživují CNS tvrdá plena (dura mater) – vystýlá lebeční a páteřní dutinu pavučnice (arachnoidea) omozečnice (pia mater) Mezi arachnoideou a pia mater je prostor vyplněný mozkomíšním mokem
Obaly CNS
Šedá hmota a bílá hmota Šedá hmota dendrity nemyelinované axony těla nervových buněk Bílá hmota axony s myelinovou pochvou Šedá hmota je i v hloubi v nižších ústředích mozku
PNS kraniální nervy – vychází z mozku a inervují hlavu a horní část těla. Je jich 12 párů. většinou obsahují sensorické i motorické neurony. některé z nich mají pouze sensorickou funkci (např. olfaktorické a optické nervy) spinální nervy – vychází z míchy a inervují celé tělo. Je jich 31 párů. Všechny obsahují sensorické i motorické neurony. ganglia
Funkční hierarchie PNS
Funkční hierarchie PNS novější dělení
PNS sensorické nervy vedou podněty z vnějšího i vnitřního prostředí do CNS motorické nervy vedou vzruchy z CNS do efektorových buněk somatický nervový systém převážně odpovědi na vnější podněty. Somatický nervový systém je většinou řízen vůlí, vědomím, ale jeho značná část pracuje reflexivně a je řízena míchou nebo nižším mozkem autonomní nervový systém
Autonomní nervový systém kontroluje vnitřní prostředí organismu: hladké svaly, srdce, trávicí soustavu, oběhovou soustavu, vylučovací soustavu tato kontrola je většinou involuntární, nevědomá autonomní systém se dělí na dvě části, které jsou mezi sebou většinou v opozici: sympatikus parasympatikus
Sympatikus a parasympatiku s
Sympatikus reakce typu „fight or flight“ (boj nebo útěk) tvorba energie, vzrušení, příprava těla pro intenzivní činnost srdce bije rychleji játra konvertují glykogen na glukosu bronchi se rozšiřují a zásobují tak plíce lépe kyslíkem trávení zastaveno stimulace sekrece adrenalinu a noradrenalinu
Parasympatikus v řadě funkcí zrcadlový opak sympatiku; konec aktivity a podpora sebeudržovacích funkcí zpomalení srdeční činnosti podpora trávení, stimulace aktivity žaludku a střev konstrikce bronchů
Autonomní nervstvo
Autonomní a somatické nervstvo …pracuje často společně např. v chladném prostředí hypotalamus signalizuje autonomnímu systému, aby zúžil průtok krve kapilárami… …a zároveň hypotalamus signalizuje somatickému nervstvu, aby způsobilo svalový třes, jehož účelem je tvorba tepla
Embryonální vývoj mozku
Embryonální vývoj mozku u všech obratlovců začíná vývoj mozku třemi oblastmi přední mozek – mnohem větší u ptáků a savců než u ryb, obojživelníků a „plazů“ střední mozek zadní mozek telencephalon u savců překryje starší oblasti mozku
Embryonální vývoj mozku u pětitýdenního embrya je již všech pět částí mozku telencephalon – vyvíjí se z předního mozku diencephalon – vyvíjí se z předního mozku mesencephalon – vyvíjí se ze středního mozku metencephalon – vyvíjí se ze zadního mozku myelencephalon – vyvíjí se ze zadního mozku
Embryonální vývoj mozku Telencephalon během druhého a třetího měsíce dochází k prudkému rozvoji telencephalonu vytváří se pravá a levá hemisféra u dospělého člověka je na povrchu kůra mozková rozbrázděná závity (gyri) obě polokoule spojuje kalozní těleso (corpus calosum)
Mozkový kmen = „nižší mozek“ je tvořen mesencephalon pons (most) z metencephalonu (zadního mozku) medulla oblongata – z myelencephalonu homeostáza organismu, kontrola pohybu, tok informací k „vyšším“ částem mozku, pomoc s komplexními pohyby, jako je chůze
Mozkový kmen („nižší mozek“) homeostáza, koordinace pohybů, vedení informací vyšším centrům medulla a pons: vysílají axony do mnoha oblasti kortexu a do mozečku (cerebellum), kde uvolňují neurotransmittery jako noradrenalin, serotonin, dopamin a acetylcholin medulla kontroluje: dýchání, aktivitu srdce a cév, polykání, zvracení, trávení na mnoha těchto aktivtách participuje též pons, např. vysílá do medully signály regulující řízení dýchání
Mozkový kmen („nižší mozek“) všechny sensorické informace, jdoucí do vyššího mozku a všechny motorické informace z vyššího mozku procházejí přes mozkový kmen rovněž integruje komplexní pohyby jako je chůze v medulle se kříží axony z vyššího mozku: levá hemisféra tak ovládá pravou stranu těla a pravá hemisféra ovládá levou stranu těla
Medulla oblongata spontánní dýchání, aktivita srdce a cév, polykání, zvracení, trávení dýchací centra kromě prodloužené míchy reguluje i pons zde v medulle dochází k překřížení axonů – levá polovina předního mozku tak odpovídá za pravou stranu těla a pravá hemisféra za levou stranu těla
Retikulární formace zatímco pons a medulla spíše odpovídají za spánek, střední mozek spíše za stav bdělosti Jedná se celkem o 90 center v mozkovém kmeni, která mají na starosti naši pozornost, stav vědomí, usínání a rovněž spánek. Retikulární formace rozhoduje o tom, jaké signály z vnějšího prostředí půjdou do vědomí a jaké ne (např. ve škole při výkladu)
Retikulární formace Neurotransmitter serotonin je produkován centry odpovídajícími za stav ospalosti. Sklenice mléka navečer může proto navodit spánek, neboť mléko obsahuje množství aminokyseliny tryptofanu, který je prekurzorem serotoninu. V cyklech bdění/spánek hraje ovšem důležitou roli hormon melatonin. Vrchol sekrece melatoninu je uprostřed noci. Jaká je funkce spánku je doposud nevysvětleno. Spánek má zřejmě vliv na paměť a učení – zdá se, že oblasti mozku, které byly aktivovány během studia jsou opět aktivovány během spánku.
Cerebellum - mozeček se vyvíjí z části zadního mozku (metencephalon) koordinuje pohyb, sensorické a kognitivní funkce pravděpodobně se podílí na učení se a zapamatování motorických dovedností, jako je např. jízda na kole atd. poškození mozečku má za následek ztrátu těchto schopností, stejně jako koordinaci ruka-oko
Diencephalon - mezimozek embryonální diencephalon se vyvíjí do tří oblastí: epithalamus, thalamus a hypothalamus epithalamus se skládá z tzv. pineální žlázy (ta produkuje hormon melatonin) a tzv. choroid plexus, což je jeden ze svazku kapilár, které produkují mozkomíšní mok. thalamus je hlavním centrem pro příchod sensorických informací a hlavním centrem pro odchod všech motorických informací z mozku. Thalamus je rovněž zapojen do prožívání emocí a vzrušení
Diencephalon - mezimozek hypothalamus sice váží jen několik gramů, avšak je nejdůležitější částí mozku pro regulaci homeostázy. obsahuje „tělní termostat“ a centra pro pocity žízně, hladu a dalších tělních funkcí hraje rovněž roli v sexuálním chování, reakcích fight or flight a v zakoušení radosti
Telencephalon – koncový mozek vyvinul se z embryonálního předního mozku, které u našich předků podporovalo především olfaktorickou (čichovou) recepci, stejně jako sluchové a optické vjemy rozdělen na pravou a levou hemisféru. V každé hemisféře je krom šedé a bílé hmoty ještě tzv. basal nuclei uloženy hluboko v bílé hmotě. Basal nuclei mají zřejmě na starost plánování a učení se pohybům (člověk s poškozenými basálními jádry není schopen poslat příkaz svalům a zůstává tak nehybný)
Telencephalon – koncový mozek nejdůležitější součástí telencephalonu je však cortex – kůra mozková. zde končí všechny sensorické informace, tvoří se motorické a vzniká řeč. Svorové těleso – corpus callosum je tvořeno silným svazkem axonů a zajišťuje komunikaci mezi oběma hemisférami
Telencephalon – koncový mozek od našich předků „plazů“ prodělal cortex výraznou expanzi u savců se část cortexu nazývá neocortex. Zatímco u krys je neocortex relativně hladký, u člověka je výrazně zbrázděný, což umožňuje, aby měl velký povrch mozek člověka má povrch 0, 5 m 2 a i když je neocortex méně než 5 mm silný, tvoří 80% celkové hmoty mozku. Krom člověka mají zbrázděný mozek ještě primáti a Cetacea, zejména velryby a delfíni cortex obsahuje 100 miliard neuronů
Cortex cortex je funkčně úzce spojen s thalamem, takže se někdy nazývá thalamocortical system skoro všechny sensorické dráhy jdoucí do cortexu prochází přes thalamus důležitou výjimkou čichové dráhy
Dominantní hemisféra je ta ve které se nachází Wernickeova oblast. U 95 % lidí je to levá hemisféra dokonce již při narození je u poloviny novorozenců Wernickeova oblast o 50 % větší v levé než adekvátní oblast v pravé hemisféře podobně Broca´s area je skoro vždy dominantní na levé hemisféře motorická oblast kontrolující ruce je u 9 lidí z 10 v levé části a většina lidí je tedy praváků
Pravá a levá hemisféra Pravá hemisféra vnímání hudby vnímání tvarů představivost vynalézavost trojrozměrná představivost kontrola pohybů levé ruky Levá hemisféra vědecké schopnosti řeč matematické schopnosti psaná řeč racionální úvahy kontrola pohybů pravé ruky
Pravá a levá hemisféra Pravá hemisféra rozeznání tváří nonverbální myšlení zpracování emocí intonace a emoční zabarvení řeči Levá hemisféra logické operace, vyžadující rychlost zpracování… …zejména v souvislosti s příkazy kosternímu svalstvu analýza zrakových a sluchových detailů Obvykle pracují obě hemisféry synchronně. Pokud má ale pacient epileptický záchvat v kalosním tělese a v levé části zorného pole vidí napsané slovo, informace nemůže přejít z pravé hemisféry do levé a pacient nemůže slovo přečíst
Pravá a levá hemisféra soustřeďte se na nos obličeje a posuďte, který z obličejů je sympatičtější pro 80 % praváků vypadá šťastněji tvář dole protože za rozeznávání a posuzování tváří je odpovědná pravá hemisféra, sledujeme levou část nakresleného obličeje obrázky nahoře i dole jsou zrcadlové obrazy …v životě je tomu zřejmě podobně: spíše si všímáme z našeho pohledu levé strany obličeje lidí (Jaynes, J. , (1990) The Origin of Consciousness in the Breakdown of the Bicameral Mind, Mariner Books, Boston)
Cortex Každá strana cortexu je dělena na čtyři laloky: frontální, temporální, okcipitální a parietální
Zpracování informací většina sensorických informací putuje skrze thalamus do příslušných korových oblastí: vizuální informace do týlního (okcipitálního) laloku akustické informace do spánkového (temporálního) laloku somatosensorické informace o doteku, bolesti, tlaku, teplotě a pozici svalů a končetin do temenního (parietálního) laloku chuťové informace do odlišných oblastí parietálního laloku olfaktorické (čichové) informace jdou nejprve do „primitivních“ oblasté kortexu a potom přes thalamus do čelního (frontálního) laloku
Primární sensorická a primární motorická oblast
Primární sensorická a primární motorická oblast místo v somatosensorické oblasti, které dostává informace např. o nohou leží těsně vedle paralelního místa motorické oblasti „mapa“ člověka neodpovídá proporcím
Jazyk a řeč francouzský lékař Pierre Broca prováděl pitvy mozků pacientů, kteří mohli rozumět jazyku, ale nemohli mluvit. Objevil že mnozí z těchto pacientů měli defekt v malé oblasti frontálního laloku (dnes zvané Brocova oblast) německý lékař Karl Wernicke nalezl poškození v zadní oblasti temporálního laloku (dnes Wernickova oblast) u pacientů, kteří mohli mluvit ale nerozuměli
Jazyk a řeč Obrázek vznikl metodou PET – pozitronová emisní tomografie: Brocova oblast je aktivní během mluvení, Wernickeova při naslouchání řeči
Jazyk a řeč
Jazyk a řeč Mozkové dráhy v situaci kdy slyším slovo a opakuji jej (nahoře) mozkové dráhy v situaci kdy slovo vidím napsané a pak jej vyslovím (dole)
Jazyk a řeč 1. 2. 3. 4. 5. 6. recepce v sluchovém ústrojí, kde se zvuky dekódují jako slova interpreetace slov ve Wernickeově oblasti determinace myšlenek vyjádřených slovy opět ve Wernickeově oblasti přenos signálů z Wernickeovy do Brocovy oblasti přes arcuate fasciculus aktivace programů v Brocově oblasti pro kontrolu tvoření slov přenos signálů do motorického cortexu které kontrolují svaly mluvidel
Nejdůležitější funkční oblasti cortexu. 95 % lidí má Wernickeovu oblast a Brocovu oblast v levé hemisféře.
Wernicke´s area somatická, visuální a auditorní oblasti se setkávají v zadní a horní části levého temporálního laloku, kde se dotýkají temporální, parietální a occipitální laloky tato oblast hraje zřejmě největší roli ve funkci mozku, nazývanou „inteligence“ oblast má mnoho různých jmen, nejznámější je jako Wernickeova oblast
Wernicke´s area při poškození Wernickeovy oblasti člověk sice slyší slova, ale nerozumí jejich významu. podobně je schopen číst, ale nerozumí významu slov elektrické dráždění oblasti, zejména pokud se elektroda dostanou hlouběji až do blízkosti thalamu, může vyvolat velmi komplexní auditorní halucinace (hudbu) nebo vynést na povrch komplikované vizuální scény z dětství (ale i dávné vzpomínky z různých sensorických zdrojů). Předpokládá se, že aktivace Wernickeovy oblasti nějak všechny tyto vzpomínky integruje, i když mohou být uloženy v různých oblastech mozku.
Čtení tzv. angular gyrus area je odpovědná za porozumění čtenému textu. Angular gyrus area leží těsně za Wernickeovou oblastí a dotýká se visuální oblasti occipitálního laloku je-li poškozena (ale Wernickeova oblast poškozena není) , osoba dobře slyší i artikuluje, ale nerozumí smyslu psaného textu porucha je zvaná dyslexia nebo word blindness pokud osoba slova slyší nebo čte ale není schopna pochopit jejich smysl, nemoc se nazývá Wernicke´s aphasia global aphasia = osoba není naprosto schopna porozumět jazyku a neí schopna komunikovat
Řeč proces řeči zahrnuje dvě stadia proces utvoření myšlenky a slova kterými má být vyjádřena motorická kontrola vokalizace samotná i zde je Wernickeova oblast velmi důležitá. Při Wernickeově afázii nebo globální afázii osoba není schopna utvořit myšlenku v mírnější podobě je schopna myšlenku utvořit ale není schopna ji vyjádřit slovy. Mluvit sice může ale proud slov nedává smysl
Motor aphasia vzniká poraněním Brocovy oblasti osoba ví co chce vyjádřit (myšlenka je přítomna), ale není schopna vydat příkaz hlasivkám svaly hrtanu, rtů, úst, respiračního systému a dalších svalů ovládajících řeč jsou ovládány z Brocovy oblasti
Wernicke´s area dlouho se mělo za to, že za „já“ či naši osobnost bude odpovědný prefrontální cortex neboť je to oblast, které je velmi výrazná u lidí ve srovnání s ostatními primáty dnes je zřejmé, že daleko důležitější oblastí je Wernicke´s area a angular gyrus
Prosophenosia spočívá v neschopnosti rozeznat tváře lidí. Dochází k ní při poškození mediálních oblastí na spodu okcipitálních laloků a medioventrálních povrchů temporálních laloků (viz obr. ) Podivuhodné je, že všechny ostatní funkce mozku jsou zcela zachovány. Je rovněž pozoruhodné, jak obrovská oblast v mozku je vyčleněna pouze pro úkol rozeznávání tváří.
Emoce celý mozek, avšak zejména limbický systém = kruhová oblast kolem mozkového kmene limbický systém se skládá: amygdala hippocampus olfactory bulb tyto struktury interagují se somatosensorickou oblastí a generují reakce jako je smích či pláč rovněž „cítění“ agrese, hladu či sexuality
Limbický systém
Limbický systém amygdala je odpovědná za rozeznání výrazů tváře druhých lidí, za rozpoznání hněvu a dobré nálady tumor či poranění těchto oblastí (kauza Phineas Gage 1848) ponechává paměť a inteligenci nedotčenu, ale změní se osobnost, motivace, rozhodovací schopnost a zaměření se na cíl
Phineas Gage
Limbický systém dříve se prováděla na zmírnění emočních poruch frontální lobotomie – přerušení spoje mezi prefrontálním kortexem a limbickým systémem. Pacienti ale vykazovali poruchy v schopnosti koncentrace, zaměřenosti na cíl proto se dnes léčí medikamenty
Myšlení, vědomí, paměť mechanismy doposud neznáme některý myšlenky vycházejí ze stimulace oblasti cortexu, thalamu, limbického systému a retikulární formace mozkového kmene teorie, která stanoví, že se všechny tyto oblasti podílí na vzniku mylšnky je zvána holistická teorie myšlení některé „hrubé“ myšlenky, jako je bolest vychází z nižších oblastí elektrická stimulace cortexu vyvolá jen mírnou bolest elektrická stimulace hypothlaamu, amygdaly a mesencephalonu vyvolá extrémně silný pocit bolesti
Paměť a učení krátkodobá paměť – čelní lalok neokortexu 7 – 10 čísel nebo znaků po dobu vteřin až minut, dokud na ně osoba myslí možná je způsobena tím, že informace krouží mezi buňkami – circuit of reverberating neurons – ale dosud se neví rovněž může být způsobena tzv. presynapticko u facilitací či inhibicí (synapse je často používána) intermediate long term memory trvá dny až týdny ale bez opakování je pak zapomenuta dlouhodobá paměť – nutná spolupráce hippocampu předpokládají se strukturální změny v synaptikcých terminálech když si po nějaké době snažíme vybavit jméno nebo číslo, z dlouhodobé paměti informaci přenášíme do krátkodobé transfer informací z krátkodobé do dlouhodobé paměti se posiluje cvičením, positivními či negativními emočními stavy z amygdaly a asociací z daty již naučenými
Negativní a pozitivní paměť kdyby si mozek pamatoval vše, co smysly zachytí, jeho kapacita by byla překročena během několika minut naštěstí má mozek schopnost ignorovat nedůležité informace, jevu se říká habituace a jedná se o typ negativní paměti pokud je vstupní informace důležitá a jejím následkem je pocit bolesti nebo radosti, paměťová stopa je uchována, jevu se říká facilitace a jedná se o typ pozitivní paměti
Habituace a facilitace u Aplysia na obrázku jsou dva synaptické terminály – první je obvyklý sensorický terminál na povrchu neuronu, který má být stimulován druhý je presynaptické zakončení ležící na povrchu sensorického terminálu a je zváno facilitatorní terminál pokud je opakovaně drážděn jen sensorický terminál, přenos signálu na postsynaptický neuron je zprvu obrovský, posléze menší a jedná se o klasickou habituaci pokud jsou drážděny oba dva terminály, signál je naopak silnější a zůstane silný po minuty, dny a až tři týdny je velmi zajímavé že i po proběhlé habituaci se drážděním obou terminálů přenos změní na facilitaci
Paměť a učení rychlé vybavení míst, telefonních čísel a faktů může spočívat v rychlých změnách síly nervových spojení pomalé učení a pamatování si dovedností (hraní tenisu) zřejmě spočívá ve vytváření zcela nových spojů mezi neurony
Na odlehčení… DOKÁŽETE PŘEČÍST NÁSLEDUJÍCÍ VĚTU? V SUOIVSOLTSI S VZÝUKEMM NA CMABRIDGE UINERVTISY VLŠYO NJAVEO, ŽE NZEÁELŽÍ NA POŘDAÍ PSÍEMN VE SOLVĚ. JEDNINÁ DLEUITŽÁ VĚC JE, ABY BLYY PNVRÍ A PSOELNDÍ PÍMESNA NA SRPVÁÉNM MSTÍĚ. ZYBETK MŽUE BÝT TOTÁNLÍ SĚMS A TY TO PŘOÁD BEZ PORLBMÉU PEŘČETŠ. JE TO PROTO, ŽE LDIKSÝ MEZOK NETČE KDAŽÉ PENSÍMO, ALE SVOLO JKAO CLEEK. ZJÍAMVAÉ, ŽE ? ? ?
Počet neuronů a jejich konektivita se výrazně mění během učení během prvních týdnů, měsíců nebo až přibližně roku života, mnoho částí mozku produkuje přebytek neuronů tyto neurony vysílají axony, které se napojují na další neurony pokud se novému axonu nezdaří se napojit na jiný neuron, sval nebo endokrinní žlázu, během týdnů se rozpouští a mizí počet nervových spojení je determinováno pomocí nerve growth factors které jsou uvolňovány ze stimulovaných buněk brzy po narození je tedy dán počet neuronů a jejich konektivita podle principu „use it or lose it“ pokud je pokusnému zvířeti po narození zakryto jedno oko na týdny, neurony degenerují a zvíře na toto oko bude buď zcela nebo částečně slepé po celý zbytek života
Počet neuronů a jejich konektivita se výrazně mění během učení i z tohoto důvodu se vřele doporučuje matkám „muchlat se“ s dětmi. . . . neboť děti tuto haptickou stimulaci životně potřebují!
Long – term potentiation presynaptický neuron uvolní krátkou salvu vysokofrekvenčních akčních potenciálů LTP trvá dny či týdny – proto je to možná fundamentální proces, kterým funguje paměť a je uskutečněno učení! neurotransmiterem je kyselina glutamová postsynaptický neuron má dva typy receptorů: AMPA a NMDA
Long – term potentiation
Anterográdní a retrodrádní amnézie anterográdní amnézie: vzpomínky z dětství a minulosti živé, v současnosti ale již pacient si není schopen nic zapamatovat dlouhodobou pamětí např. po odstranění hippocampu retrográdní amnézie: pacient si není schopen vybavit vzpomínky z mládí, ale pro přítomné události paměť funguje dobře např. při poškození thalamu, který zřejmě nějak pomáhá „vykopávat“ vzpomínky z minulosti
Vědomí zcela jistě přesahuje do filosofie a náboženství pomocí magnetické rezonance se srovnávají stavy mozku při různých stavech vědomí – „centrum vědomí“ v mozku zřejmě neexistuje – spíše se zdá že mozek umí opakovaně „skenovat“ sám sebe a že vědomí je výslednicí různých aktivit do jednoho obrazu vztah mezi vědomím a neurony je možná podobný hurikánu a jednotlivým molekulám vody
Celkový přehled