Strategie regulace proteinov enzymov aktivity 1 Allosterick kontrola
 enzymové aktivity. 1. Allosterická kontrola. Allosterické proteiny obsahují regulační místa odlišná")


. ATCasa katalyzuje kritický stupeň")
 inhibuje ATCasu. Podobnost mezi substrátem a produktem není žádná.")


 Sedimentační rychlost nativní ATCasy. B) Enyzm po působení p-hydroxymerkuribenzoanu. Enzym")
.")
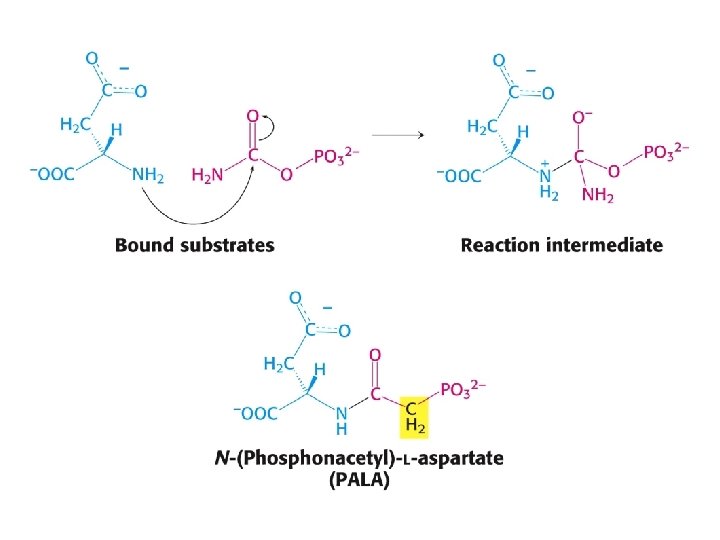
-L-aspartát.")

 stabilizuje T stav. CTP se váže na regulační podjednotky ATCasy.")









 model – kvantitativní formulace. • Mějme enzym s n aktivními")




 na podjednotky proteinu vede")

 a John Kendrew – Nobelova cena za chemii v")









 na afinitu kyslíku k hemoglobinu. • Bez")
 na lidský hemoglobin. Váže se do centrální dutiny")













. • • Laktátdehydrogenasa Isoenzymy H a M")













 aktivuje proteinkinasu A změnou její struktury.")



 buňkami pankreatu.")

 a chymotrypsinu (modře). • Elektrostatická interakce mezi karboxylem Asp 194 a")


, inhibitor (červený). Lys 15 inhibitoru vstupuje do")



, also known as the Williams-Fitzgerald-Flaujeac factor or the")
 štěpí fibrinopeptidy z centrální globule fibrinogenu. Vytvořené peptidy")
.")

. Působením antagonistů vit. K")

 není v krvi přítomen antihemofilní faktor VIII nebo")



- Slides: 95
Strategie regulace (proteinové) enzymové aktivity. 1. Allosterická kontrola. Allosterické proteiny obsahují regulační místa odlišná od substrátových. Allosterický z řečtiny „allos“ = další, „steros“ = uspořádání. Skládají se z podjednotek (protomerů). Mají schopnost kooperativity. 2. Jako příklady enzym aspartáttranskarbamoylasa (ATCasa) a neenzymový kyslíkový přenašeč hemoglobin. 3. 2. Mnohočetné formy enzymů. Isozymy – jsou homologní enzymy katalyzující stejnou reakci, ale lišící se jemně v struktuře a více v Km a Vlim a regulačních vlastnostech.
• 3. Reversibilní kovalentní modifikace. Katalytické vlastnosti řady enzymů se mění po kovalentní vazbě nějaké skupiny na jejich molekulu – nejčastěji fosforylace. Modifikující enzymy jsou proteinkinasy a proteinfosfatasy. • 4. Proteolytická aktivace. Mnoho proteinů je syntetizováno v neaktivní formě. U enzymů jsou to zymogeny (proenzymy). Po odštěpení části řetězce přechází zymogen na aktivní enzym. Jako aktivující složky působí proteolytické enzymy jako chymotrypsin, pepsin a trypsin. Aktivace je ireversibilní. Kaspasy, proteolytické enzymy, účastnící se programové buněčné smrti, jsou aktivovány z formy prokaspas. Dalším příkladem je kaskáda enzymů při srážení krve.
ALLOSTERIE • Aspartáttranskarbamoylasa jako příklad allosterického enzymu. • Allosterické enzymy se neřídí kinetikou Michaelise a Mentenové. • Modely allosterie: • Symetrický model ( Jacques Monod, Jeffries Wyman a Jean. Pierre Changeux – podle autorů zkracovaný jako MWC model). • Sekvenční model (Daniel Koshland). • Conceptual insights – Stryer, Chapter 10. • Hemoglobin jako příklad allosterického proteinu
Aspartáttranskarbamoylasa je allostericky inhibována konečným produktem metabolické cesty cytidintrifosfátem (CTP). ATCasa katalyzuje kritický stupeň kondnzace Asp s karbamoylfosfátem při syntéze pyrimidinových nukleotidů.
Cytidintrifosfát (CTP) inhibuje ATCasu. Podobnost mezi substrátem a produktem není žádná.
Od slide 6 po slide 13 je prezentován způsob objasnění struktury ATCasy. Není podstané pro pochopení principu allosterie !! Modifikace cysteinových vedlejších řetězců (ATCasy).
Centrifugační studie ATCasy. • Sedimentační koeficient nativního enzymu je 11, 6 S. • Dvě podjednotky 2, 8 S a 5, 8 S. • Jak od sebe oddělíme obě podjednotky a jak lze odstranit p-hydroxymerkuribenzoát ? • Větší podjednotka (c) je katalytická, ale není inhibována CTP. Složena ze tří řetězců po 34 k. D se označuje c 3. • Regulační podjednotka (r) váže CTP. Složena ze dvou řetězců po 17 k. D je označena r 2. • Nativní enzym je složen ze dvou katalytických trimerů a tří regulačních dimerů.
Ultracentrifugační studie ATCasy. A) Sedimentační rychlost nativní ATCasy. B) Enyzm po působení p-hydroxymerkuribenzoanu. Enzym disocioval na regulační (r) a katalytické podjednotky (c).
Kvarterní struktura ATCasy. Vpravo je zjednodušený pohled. Katalytické trimery jsou dva (druhý je zakrytý). Regulační dimery jsou tři. 2 c 3 + 3 r 2 = c 6 r 6
PALA – bisubstrátový analog reakčního meziproduktu. PALA = N-(fosfonoacetyl)-L-aspartát.
Aktivní místo ATCasy. V aktivním místě, které obsahuje hlavně vedlejší skupiny z jedné podjednotky, ale také část druhé podjednotky (zeleně), je vázán PALA.
Cytidintrifosfát (CTP) stabilizuje T stav. CTP se váže na regulační podjednotky ATCasy.
T a R stavy v rovnováze. I v situaci bez přítomnosti substrátu existuje ATCasa v rovnováze obou stavů. Bez substrátu převažuje T nad R stavem faktorem 200.
ATCasa vykazuje sigmoidní kinetiku.
Tvorbu sigmoidní křivky lze interpretovat jako směs dvou Michaelis-Mentenovských enzymů, jeden s vysokou hodnotou KM odpovídající T stavu a druhý s nízkou hodnotou KM odpovídající R stavu. S růstem koncentrací substrátu se rovnováha posunuje od T k R stavu.
Vliv CTP na kinetiku ATCasy. CTP je allosterický inhibitor, stabilizuje T stav. Znesnadňuje vstup substrátu do aktivního místa.
Vliv ATP na kinetiku ATCasy. ATP je allosterický aktivátor. Stabilizuje stav R – usnadněný vstup substrátu do aktivního místa.
• Působení substrátů na allosterické enzymy se označuje jako homotropní efekt (z řečtiny „stejný“). • Působení nesubstrátových molekul na allosterické enzymy jako např. CTP a ATP na ATCasu se označuje jako heterotropní efekt. • Zkuste uvažovat proč působí ATP jako aktivátor ATCasy (fyziologická úvaha).
Dva modely allosterie. • Symetrický model ( Jacques Monod, Jeffries Wyman a Jean-Pierre Changeux – podle autorů zkracovaný jako MWC model). • Sekvenční model (Daniel Koshland).
• Conceptual insights – Chapter 10.
Kvantitativní popis modelu MWC. Frakční aktivita Yf je část aktivních míst s navázaným substrátem úměrná reakční rychlosti. a je poměr [ S ] / disociační konstantě S s enzymem ve stavu R; L je poměr koncentrací enzymu ve stavu T k enzymu ve stavu R. Vazba regulátorů ATP a CTP mění hodnotu L.
• Symetrický (MWC) model – kvantitativní formulace. • Mějme enzym s n aktivními místy. • Dva stavy enzymu v rovnováze: R – vysoká afinita k substrátu, T - nízká afinita k substrátu. Definujeme c jako poměr afinit obou forem enzymu k substrátu S, měřeno jako disociační konstanty.
• Frakční aktivita Y, je frakce aktivních míst s navázanými substráty. Je přímo úměrná reakční rychlosti. a je poměr [ S ] ku disociační konstantě S v komplexu s enzymem v R stavu. • L je poměr koncentrace enzymu v T stavu ku koncentraci enzymu v R stavu (KR). • Vazba regulátorů jako je ATP nebo CTP na ATCasu mění hodnotu L a tím odezvu na koncentraci substrátu.
Příklad využití kvantitativního modelu s daty ATCasy. • U ATCasy se n = 6. • Vynikajícího souhlasu s experimentálními daty se dosahuje při L = 200 a c = 0, 1. • Z toho plyne, že za absence vázaného substrátu je posunuta rovnováha na stranu T faktorem 200 ( pouze 1 molekula z 200 je ve stavu R) a afinita R stavu k substrátu je 10 x vyšší než ke stavu T. Když se substrát váže do každého aktivního místa, rovnováha se posouvá k R. • Např. : Pokud je obsazena polovina aktivních míst substrátem(tři ze šesti) je poměr T/R 1 : 5. Znamená to, že téměř všechny molekuly jsou ve stavu R.
• Vliv CTP a ATP se řeší jednoduše změnou hodnoty L. • Při saturaci CTP se hodnota L zvyšuje na 1 250. To znamená, že je třeba daleko více substrátu k posunu rovnováhy do stavu R. Při saturaci ATP klesá hodnota L na 70.
SEKVENČNÍ MODEL • Daniel KOSHLAND – postupná vazba ligandů (substrátů) na podjednotky proteinu vede k postupnému vzájemnému ovlivňování jednotlivých podjednotek. • Možno vysvětlit i negativní kooperaci. • U symetrického modelu (MWC) přechází celý enzym z jednoho stavu do druhého (T a R).
Jednoduchý sekvenční model tetramerního allosterického enzymu. Postupnou vazbou substrátu na podjednotky se mění konformace okolních ze stavu T na R.
Max Perutz (1914 – 2002) a John Kendrew – Nobelova cena za chemii v roce 1962. • Max Perutz pracoval na rentgenové struktuře Hb více než 30 let na Univerzitě v Cambridge (UK). Struktura o rozlišení 2, 8 A byla získána až v roce 1968. • John Kendrew navazoval na M. Perutze, Objasnil strukturu velrybího myoglobinu – rentgenostrukturní analýzou, metodou isomorfní záměny. Těžký atom jako Hg 2+ (bohatý na elektrony) se musí vázat na protein, tak, aby nedošlo ke změně struktury. Dochází k podstatné změně v intenzitě odrazu, ale ne v pozici elektronové hustoty.
Kvartérní struktura hemoglobinu – dva řetězce a a dva b tvoří pár ab dimerů – heteroteramer a 2 b 2.
Kooperativní zvýšení transportu kyslíku u hemoglobinu. Kooperativita mezi vazebnými místy pro kyslík vede k zásobení tkání větším množstvím kyslíku (1, 7 x) než by mohl nekooperativní protein. (p. O 2 je parciální tlak kyslíku)
Krevní oběh – srdce a plíce. Formy Hb: Hb. O 2 = oxyhemoglobin, Hb=deoxyhemoglobin, Mb = svalový myoglobin.
Pozice Fe 2+ v deoxyhemoglobinu.
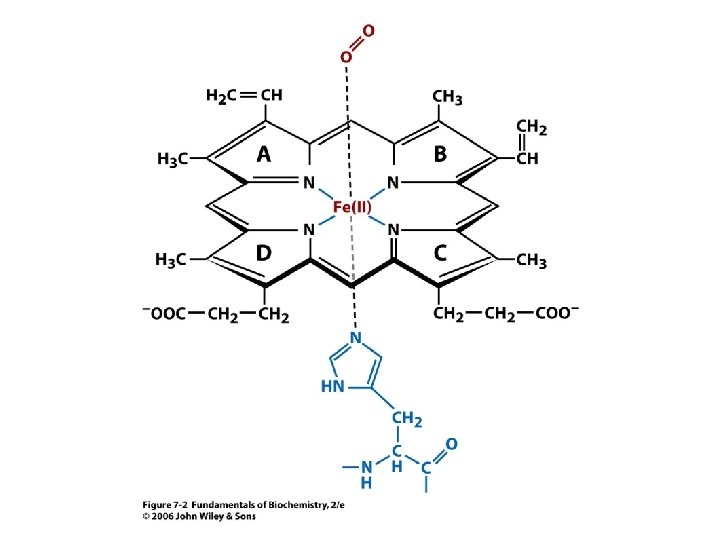
Vazba kyslíku vede ke strukturním změnám. Atom Fe je vtažen po vazbě kyslíku do roviny hemu. S ním je vtažen i proximální His.
Přechod ze stavu T do stavu R u hemoglobinu. Po vazbě kyslíku na pár a 1 b 1 se a 1 b 1 pár pootočí vůči druhému a 2 b 2 páru o 15 o.
Posun mezi řetězci b 2 a a 1 hemoglobinu po vazbě kyslíku. Stavy R vázán kyslík a T bez kyslíku.
Konformační změny v hemoglobinu. Pohyb Fe 2+ po vazbě kyslíku vtáhne do kruhu proximální His, který jako součást a helixu ovlivní svým C koncem vztah mezi ab páry vedoucí k dalším strukturním změnám. Porovnáni se strukturou deoxy (šedá).
Který z modelů je ten, který lze aplikovat na kooperativní vazbu kyslíku na hemoglobin (Hb) ? • Žádný z obou modelů nelze na Hb čistě aplikovat. • Vyhovuje spíše kombinace obou modelů. • Hb se třemi místy obsazenými kyslíkem splňuje kriteria sekvenčního modelu – nachází se v R stavu. Zbývající čtvrté vazebné místo má afinitu ke kyslíku 20 x vyšší než plně deoxygenovaný Hb vážící svůj první kyslík. Kriteria sekvenčního modelu však nejsou splněna, protože Hb s kyslíkem navázaným pouze v jednom místě ze čtyř zůstává v T stavu. Tato struktura váže kyslík 3 x pevněji než plně deoxygenovaný Hb – což odpovídá sekvenčnímu modelu. • Závěr: Oba modely reprezentují idealizované limitní stavy, kterým může reálný vyhovovat, ale častěji nevyhovuje.
Vliv 2, 3 -bisfosfoglycerátu (2, 3 -BPG) na afinitu kyslíku k hemoglobinu. • Bez přítomnosti 2, 3 -BPG je afinita hemoglobinu ke kyslíku podstatně vyšší a uvolňuje se jen 8 % přenášeného kyslíku do tkání. • 2, 3 -DPG je přítomen v erythrocytech v koncentraci 2 m. M (stejná jako konc. hemoglobinu).
Způsob vazby 2, 3 -BPG (-5) na lidský hemoglobin. Váže se do centrální dutiny deoxyhemoglobinu na osm kladně nabitých skupin obou b řetězců.
Proč 2, 3 -BPG tak silně ovlivňuje afinitu hemoglobinu ke kyslíku ? • 2, 3 -BPG se váže přednostně na deoxyhemoglobin, který stabilizuje. Deoxyhemoglobin je ve stavu T. Přechod do stavu R se děje jen po oddělení 2, 3 -BPG ! • Fyziologický důsledek: Plodový (fetální) hemoglobin se liší od hemoglobinu dospělého člověka. Je to tetramer obsahující dva řetězce a a dva g !! Řetězce g jsou ze 72 % identické s řetězci b u hemoglobinu dospělých. Tato změna však znamená, že řetězce g mají o dva pozitivní náboje méně než b a tím je podstatně snížena afinita fetálního hemoglobinu k 2, 3 -BPG a tím zvýšena afinita ke kyslíku ve srovnání s hemoglobinem mateřským. • To umožňuje bezproblémový přenos kyslíky z matky na plod.
Afinita ke kyslíku u fetálních erythrocytů.
Bohrův efekt. • Rychle metabolizující tkáně, jako svalstvo, mají velké požadavky na kyslík a produkují velké množství H+ a CO 2. Jak H+, tak CO 2 jsou heterotropní efektory hemoglobinu zvyšující uvolňování kyslíku. Afinita hemoglobinu ke kyslíku klesá při vstupu hemoglobinu do oblastí s nižším p. H. • Transport z plic (p. H 7, 4), parciální tlak kyslíku 100 torrů do aktivního svalu s p. H 7, 2 a parciálního tlaku kyslíku 20 torrů vede k uvolnění 77 % celkové přenášené kapacity. • Pokud by nedošlo k poklesu p. H, tak by se uvolnilo pouze 66 %. • Ve tkáních, kde je vyšší koncentrace CO 2 klesá afinita hemoglobinu ke kyslíku. • Za přítomnosti CO 2 o parciálním tlaku 40 torrů se uvolní z hemoglobinu 90 % přenášeného kyslíku. • Vliv H+ a CO 2 na regulaci vazby kyslíku na hemoglobin se nazývá Bohrův efekt. Podle Christiana Bohra, který jev popsal v roce 1904.
Vliv p. H a koncentrace CO 2 na afinitu kyslíku k hemoglobinu.
Chemický základ Bohrova efektu. • V deoxyhemoglobinu tvoří tři aminokyseliny dvě iontové vazby stabilizující T stav. Tvorba jedné z vazeb závisí na přítomnosti dalšího protonu na His b 146. To umožňuje vazbu na Asp 94.
CO 2 stabilizuje deoxyhemoglobin reakcí s terminální aminoskupinou za tvorby karbamátu. Změna náboje vede k pevnější vazbě mezi ab dimery a tím ke stabilizaci T stavu.
• Hemoglobin s navázanými CO 2 a H+ vstupuje do plic, kde uvolní CO 2 a H+ a naváže kyslík. • Hemoglobin se však podílí pouze 14 % na celkovém transportu oxidu uhličitého a protonů. • Jak CO 2, tak H+ jsou transportovány krví jako hydrogenuhličitan (HCO 3 -), který se tvoří spontánně nebo enzymově karbonátanhydrasou.
MYOGLOBIN
Vazba kyslíku na myoglobin – myoglobin je zpoloviny nasycen kyslíkem při tlaku kyslíku (p. O 2) 2, 8 torru.
Fyziologická role myoglobinu. • • Rolí myoglobinu je usnadňovat difůzi kyslíku ve svalech při intenzívní svalové činnosti. Rychlost difůze kyslíku z krevních kapilár do tkání je limitována jeho nízkou rozpustností ve vodě (10 -4 M v krvi). Myoglobin zvyšuje efekt rozpustnosti kyslíku ve svalových buňkách. Myoglobin jako „sklad“ kyslíku je důležitý u mořských savců jako je např. velryba, jejichž koncentrace myoglobinu ve svalech je 10 x vyšší než u suchozemských savců. Myši u kterých byl vyřazen („knocked out“) gen pro myoglobin, mají světlejší svalstvo, ale jinak jsou normální. Myoglobin není pro život suchozemských savců nezbytný. V mozku byl nedávno popsán protein charakteru myoglobinu nazvaný neuroglobin. Funkcí je podpora koncentrace O 2 v nervových tkáních, které jsou metabolicky velmi aktivní. Mozek reprezentuje asi 2% hmotnosti lidského těla, ale spotřebovává 20% získaného kyslíku.
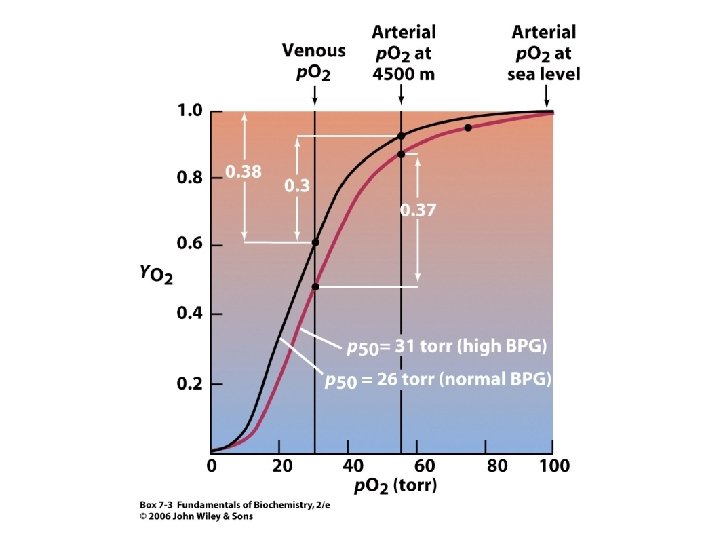
Adaptace na výšku. • • Atmosférický tlak klesá s výškou. Tlak kyslíku ve výšce 3 000 m je 110 torrů, tj. 70 % hodnoty tlaku na hladině moře. Tlak kyslíku pod 85 torrů je neslučitelný se životem. Adaptace na výšku je spojena se zvýšením množství Hb v erythrocytech a zvýšením počtu erythrocytů. Tento komplexní proces obvykle zahrnuje několik týdnů. Mnohdy stačí pouze jeden k dosažení dostatečné adaptace. Tento efekt spočívá v rapidním zvýšení množství BPG syntetizovaného v erythrocytech (obvykle ze 4 m. M na 8 m. M; BPG nemůže procházet membránou erythrocytu). Zvýšená koncentrace BPG způsobuje, že se křivka saturace Hb kyslíkem posunuje oproti křivce na hladině moře do pozice znamenající nižší afinitu !! Na hladině moře je rozdíl p. O 2 mezi tepennou a žilní krví 70 torrů a Hb uvolňuje 38 % vázaného kyslíku. Když obsah vázaného kyslíku v tepnách klesne na 55 torrů, to je ve výšce 4 500 m, uvolňuje Hb jen 30 % vázaného kyslíku. Adaptace na výšku (pokles vázaného kyslíku v plících, zvýšení množství kyslíku uvolněného v tkáních) znamená, že Hb přenáší prakticky normální množství kyslíku.
• Koncentrace BPG se zvyšuje v erythrocytech mnohem rychleji než proběhne syntéza Hb (erythrocyty nemají jádro a proto nemohou syntetizovat proteiny). • Hladina BPG je mnohem citlivější regulace přenosu kyslíku než změna respirační rychlosti. • Hyperventilace, další rychlá odezva na výšku, vede k respirační alkalose. • Alkalosa (p. H větší než 7, 6) a acidosa (p. H menší než 7, 1) jsou následky nerovnováhy krevních ústojů. • Hyperventilace urychluje vydechování CO 2 a způsobuje respirační alkalosu. • Čím je způsobována acidosa a jak se dá potlačit ? ? • Alkalosa se dá potlačit Na. Cl nebo KCl (proč? ) nebo vdechováním vzduchu obohaceneého CO 2.
• Obyvatelé And a Himalájí mají vysokou kapacitu plic, vysokou hladinu Hb a často také větší pravou srdeční komoru což znamená zvýšení objemu čerpané krve. • Na rozdíl od adaptace lidí na výšku, většina savců žijících ve vysokých výškách (lamy), má geneticky pozměněný Hb vyznačující se vyšší afinitou ke kyslíku.
2. Mnohočetné formy enzymů - isozymy (isoenzymy). • • Laktátdehydrogenasa Isoenzymy H a M Interakce isoenzymů LDH se substráty a inhibitory. Isoenzymy LDH v různých fázích vývoje organismu a v různých orgánech
ISOENZYMY – způsob regulace v různých tkáních a za různých vývojových stádií. • Isozymy nebo isoenzymy jsou enzymy lišící se sekvencí aminokyselin katalyzující stejnou reakci. • Obvykle mají různé kinetické parametry, jako KM, nebo různé aktivátory a inhibitory. • Isoenzymy od sebe obvykle odlišíme dle jejich různé elektroforetické pohyblivosti. • Jako typický příklad lze uvést isoenzymy laktátdehydrogenasy (LDH). LDH se uplatňuje při anaerobní metabolismu glukosy a syntéze glukosy (glukoneogeneze). • U člověka rozeznáváme dva isoenzymové polypeptidové řetězce: H isoenzym – vysoký obsah v srdečním svalu a M isoenzym – skeletální svaly. Řetězce jsou ze 75 % shodné. • Funkční enzym je tetramer.
• H 4 isoenzym je hojný v srdečním svalu a má větší afinitu k substrátům než M 4 isoenzym. • Vysoká hladina pyruvátu allostericky inhibuje H 4 a ne M 4. Proč ? ? • Další kombinace, jako je H 3 M, mají vlastnosti mezi H 4 a M 4. • M 4 isoenzym je optimální za anaerobních podmínek, H 4, za aerobních. • Přítomnost různých kombinací isoenzymů se mění s vývojem např. srdce. • Přítomnost různých isoenzymů v krvi je indikátorem poškození tkáně a tento fakt se využívá k diagnostickým účelům. • Např. zvyšující se hladina H 4 v séru ve vztahu k H 3 M je indikací infarktu myokardu.
Isoenzymy LDH – krysí srdeční sval. • Krysí srdeční LDH isoenzym. Profil se mění s vývojem. • H isoenzymy jsou modré kroužky, M jsou červené čtverce.
Obsah LDH isoenzymů v různých tkáních krysy.
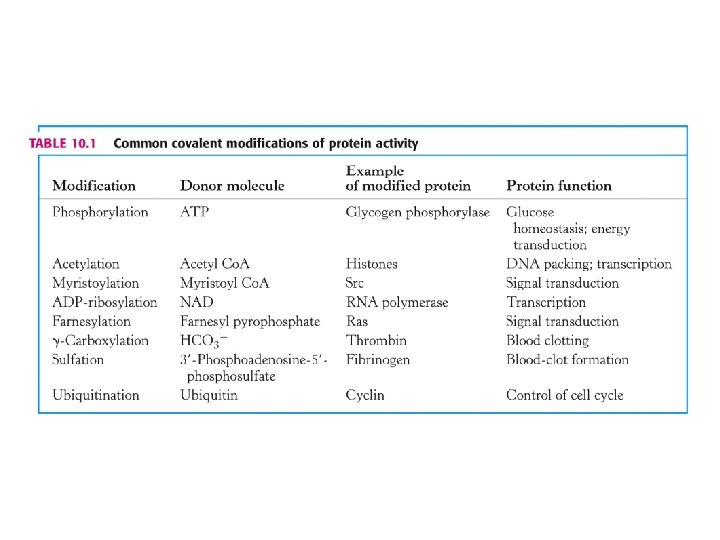
3. Regulace enzymové aktivity kovalentní modifikací. • Nejčastější jsou fosforylace a defosforylace • Dalším způsobem modifikace je např. acetylace. Acetylovány jsou histony (pomocné proteiny obalující DNA v chromosomech a při regulaci genů). Vysoce acetylované histony spojené s geny jsou aktivně přepisovány. Enzymy jsou acetyltransferasa a deacetylasa a jsou regulovány fosforylací. • Modifikace není vždy reversibilní. Např. připojení ubiquitinu k proteinu.
Kovalentní modifikace proteinů – acetylace vedlejšího řetězce Lys
Vysvětlivky k tabulce: Common covalent modification of protein activity • Ras = onkogen – rat sarcoma viruses. • Src = onkogen – Rous sarcoma viruses. • Myristoyl-Co. A = CH 3 – (CH 2)12 – CO-Co. A • Farnesylpyrofosfát (3 x prenyl, prenylace) –
Fosforylace je vysoce účinný způsob modifikace regulující aktivitu cílového proteinu.
Tabulka několika známých proteinkinas.
Proteinfosfatasy obrací působení proteinkinas.
Fosforylace a defosforylace nejsou procesy samozřejmně reversibilní. Každý je za fyziologických podmínek ireversibilní. Probíhají pouze za přítomnosti enzymů. • Rychlost přechodu fosforylované a nefosforylované formy závisí na relativní aktivitě kinasy a fosfatasy. • Výsledek dvou reakcí závisí na hydrolýze ATP na ADP a Pi (- 50 k. J/mol za podmínek buňky).
Proč je fosforylace tak efektívní způsob regulace aktivity proteinů ? • 1. Fosfát vnáší do molekuly proteinu dva negatívní náboje. Původní elektrostatické interakce jsou rušeny a vznikají nové. Modifikuje vazbu substrátu atp. • 2. Fosfátová skupina může tvořit tři nebo více vodíkových vazeb. Tetrahedrální geometrie fosfátových skupin dává těmto vodíkovým vazbám jednoznačný směr. • 3. Volná energie fosforylace je vysoká (- 50 k. J/mol). Fosforylace tak posunuje konformační rovnováhu mezi různými funkčními stavy faktorem 104.
• 4. Fosforylace a defosforylace probíhají v časech kratších než sekunda, maximálně v hodinách. Reaguje se tak rychle na fyziologickou potřebu. • 5. Fosforylace obvykle vede k podstatnému zvýšení rychlosti. Kinasa rychle fosforyluje velké množství proteinů. • 6. ATP je energetické platidlo buňky. Použití ATP je spojeno s energetickým stavem buňky.
Cyklický AMP (c. AMP) aktivuje proteinkinasu A změnou její struktury.
Regulace proteinkinasy A. Vazbou čtyř molekul c. AMP je proteinkinasa A aktivována. Inhibovaný holoenzym (R 2 C 2) disociuje po vazbě c. AMP na regulační podjednotku (R 2) a katalyticky aktivní podjednotku ©.
4. Mnoho enzymů je aktivováno specifickým proteolytickým štěpením.
• Specifická proteolýza: • 1. Trávící enzymy jsou syntetizovány jako zymogeny v žaludku a pankreatu. • 2. Srážení krve je modifikováno kaskádou proteolytických aktivací, které jsou rychlou odezvou na poranění. • 3. Některé proteinové hormony jsou syntetizovány jako inaktivní prekurzory. Např. insulin je syntetizován jako preproinsulin, proteolyticky se oddělují dva peptidy. • 4. Fibrilární protein kolagen, hlavní součást kůže a kloubů, je tvořen z prokolagenu, rozpustného prekurzoru. • 5. Mnohé vývojové procesy jako např. metamorfóza pulce na žábu. Velké množství kolagenu je resorbováno z bičíku v několika dnech. • 6. Programová smrt buňky, apoptóza, je zprostředkována proteázami jako je kaspasa, syntetizované z prekurzoru prokaspasy.
Sekrece zymogenů acinárními (hroznovitými) buňkami pankreatu.
Proteolytická aktivace chymotrypsinogenu. Tři řetězce a-chymotrypsinu jsou vázány dvěma disulfidovými vazbami mezi řetězci (A k B a B k C).
Konformace chymotrypsinogenu (červeně) a chymotrypsinu (modře). • Elektrostatická interakce mezi karboxylem Asp 194 a a-aminoskupinou Ile 16 je nutná pro strukturu aktivního chymotrypsinu.
Aktivace zymogenů proteolytickým štěpením. Aktívní enzymy jsou žlutě, zymogeny oranžově. Enteropeptidasa iniciuje aktivaci. Trypsin je společný aktivátor všech pankreatických zymogenů. Buňky dvanácterníku vylučují enteropeptidasu, která hydrolyzuje Lys-Ile vazbu trypsinogenu.
Inhibitory proteolytických enzymů. • Aktivace zymogenů je ireversibilní. K omezení nebo inaktivaci aktivních enzymů se využívají specifické inhibitory proteas. • Např. pankreatický trypsinový inhibitor je 6 kd protein inhibuje trypsin tak, že se váže velmi pevně do jeho aktivního místa. Inhibiční konstanta je 1 p. M !! • Komplex nelze oddělit ani denaturací 8 M močovinou nebo 6 M guanidin. HCl.
Interakce trypsinu se svým inhibitorem. Trypsin (žlutý), inhibitor (červený). Lys 15 inhibitoru vstupuje do aktivního místa. Vytváří iontovou vazbu s Asp 189.
Proč existuje trypsinový inhibitor ? ? • Trypsin aktivuje celou řadu zymogenů. Inhibitorem je inaktivována jen velmi malá část trypsinové aktivity. • Molekuly trypsinu aktivované ve žlučníku nebo v pankreatu by způsobily poškození tkáně – akutní pankreatitida.
Srážení krve se uskutečňuje kaskádou aktivace zymogenů. • Sraženina fibrinu se tvoří souhrou vnitřních a vnějších faktorů a v konečné fázi společných drah. • Vnitřní f. začíná aktivací faktoru XII kontaktem s neobvyklým povrchem způsobeným zraněním. • Vnitřním faktorem je trauma aktivující faktor VII a uvolňující lipoprotein z krevních destiček. • Inaktivní srážecí faktor je červeně, aktivovaný žlutý. Stimulační proteiny jsou modře. • Principem je, že aktivovaný faktor aktivuje faktor následující.
Kaskáda srážení krve – kaskáda aktivací zymogenů. Fibrin se tvoří souhrou vnitřních, vnějších a v konečné fázi společných drah. Vnější – poranění, vnitřní trauma po poranění. XII- Hagemanův faktor, „tissue factor“ = lipoprotein z krevních destiček
Kininogen, kallikrein. • High-molecular-weight kininogen (HMWK), also known as the Williams-Fitzgerald-Flaujeac factor or the Fitzgerald factor or the HMWK-kallikrein factor, is a protein from the blood coagulation system as well as the kinin-kallikrein system. It is a protein that adsorbs to the surface of biomaterials that come in contact with blood in vivo. This protein circulates throughout the blood and quickly adsorbs to the material surfaces.
Tvorba fibrinové sraženiny. Thrombin (proteolytický enzym) štěpí fibrinopeptidy z centrální globule fibrinogenu. Vytvořené peptidy polymerují na způsob sítě.
Nově vytvořená sraženina se stabilizuje zesíťováním za účasti transglutaminasy (faktor XIIIa).
Thrombin je syntetizován ze zymogenu - prothrombinu. • • • Prothrombin obsahuje čtyři domény. Aktivace se uskutečňuje proteolytickým štěpením. Vitamin K je nutný k syntéze prothrombinu. Antagonisté vitaminu K jsou dikumarol a warfarin. Dikumarol se používá v lékařství jako antikoagulant. Warfarin a další antagonisté vit. K slouží jako jed na hlodavce. • Za přítomnosti antagonistů vitaminu K je syntetizován abnormální prothrombin, který neváže Ca 2+.
Normální prothrombin váže Ca 2+ modifikovanou aminokyselinou g-karboxyglutamátem (silná chelatace). Působením antagonistů vit. K dochází k syntéze prothrombinu bez vázaného Ca 2+ • Vazba Ca 2+ umožňuje prothrombinu (zymogenu) vstoupit do fosfolipidové membrány vzniklé z krevních destiček po poranění. Tím se dostává do blízkosti proteinů, které ho aktivují na thrombin.
Úloha vitaminu K a jeho antagonistů při srážení krve. • Vitamin K je nutný k syntéze prothrombinu a dalších faktorů srážení krve. • Antagonisty vitaminu K jsou dikumarol a warfarin. • Oba antagonisté se používají jako antikoagulancia.
Hemofilie. Při klasické hemofilii (dědičná, A) není v krvi přítomen antihemofilní faktor VIII nebo je jeho aktivita nízká. Antihemofilní faktor (VIII) – stimuluje aktivaci faktoru X faktorem IXa. Léčí se transfůzí frakce koncentrované plasmy obsahující VIII.
K zamyšlení. • • 1. His v aktivním místě aspartáttranskarbamoylasy je nutný ke stabilizaci přechodového stavu vázaných substrátů. Jaká bude závislost katalytické rychlosti na p. H ? Uvažujte, že tato reakce je dominantní pro p. H profil enzymu. 2. Představte si, že jste vyizolovali dimerní enzym obsahující dvě stejná aktivní místa. Vazba substrátu do jednoho aktivního místa snižuje afinitu k substrátu druhého aktivního místa. Který allosterický model nejlépe vyjadřuje tuto negativní kooperativitu ? 3. Erythrocyty ptáků a želv obashují regulační molekuly odlišné od 2, 3 -BPG. Tyto látky jsou také účinné při snižování afinity hemoglobinu ke kyslíku, dokonce i lidského hemoglobinu zbaveného 2, 3 -BPG. Která z následujících látek by mohl být v tomto smyslu nejúčinnější ? a) Glukosa-6 -fosfát; b) inositolhexafosfát; c) HPO 42 - ; d) Malonát; e) Arginin; f) Laktát.
• • • 4. Prostředí ovlivňuje částečně p. K kyseliny. Jaký vliv budou mít následující změny prostředí na p. K vedlejší řetězce Glu ? a) Do blízkosti se dostane vedlejší řetězec Lys. b) Do blízkosti se dostane terminální karboxyl proteinu. c) Vedlejší řetězec Glu je posunut z vnějšku proteinu do hydrofobní kapsy uvnitř. 5. Jaký vliv má každá následující změna na afinitu hemoglobinu ke kyslíku in vitro ? a) Zvýšení p. H ze 7, 2 na 7, 4; b) Zvýšení p. CO 2 z 10 na 40 torrů; c) Zvýšení [2, 3 -BPG] z 0, 2 na 0, 8 m. M; d) Disociace a 2 b 2 na monomerní podjednotky. 6. Substrát se váže 100 x pevněji na R stav než na T stav allosterického enzymu. Aplikuj na enzym model MWC. a) Jakým faktorem posune vazba jedné molekuly substrátu na enzym poměr koncentrací enzymových molekul v R a T stavech ? b) Uvažujte že, poměr [T] /[R] za nepřítomnosti substrátu je 107 a, že enzym obsahuje čtyři vazebná místa pro substrát. Jaký je poměr enzymových molekul ve stavu R ku stavu T za přítomnosti saturačního množství substrátu ? 7. Allosterický enzym vyhovující modelu WMC má poměr T/R = 300 bez přítomnosti substrátu. Uvažujte mutaci, která poměr obrátí. Jak může taková mutace ovlivnit vztah mezi rychlostí reakce a koncentrací substrátu?
• • • Odpovědi: 1. Protonizovaná forma His pravděpodobně stabilizuje negativní náboj kyslíkového atomu štěpitelné vazby v přechodovém stavu. Deprotonizace by mohla vést ke snížení aktivity. Lze uvažovat, že rychlost bude polovina maximální při p. H kolem 6, 5 (p. K His vedlejšího řetězce v proteinu) a poklesne při zvýšení p. H. 2. Sekvenční model, model MWC ne. 3. b) Inositolhexafosfát. 4. Hodnota p. K se a) snižuje, b) zvyšuje a c) zvyšuje. 5. a) zvyšuje se, b) snižuje se, c) snižuje se a d) zvyšuje se afinita ke kyslíku. 6. a) 100, změna poměru [R]/[T] po vazbě jednoho substrátu musí být shodná s poměrem substrátových afinit obou forem. b) 10, vazba čtyř molekul substrátu mění poměr [R]/[T] faktorem 1004 = 108. Poměr za nepřítomnosti substrátu je 10 -7. Z toho plyne, že poměr v plně nasycené molekule je 108 x 10 -7 = 10. 7. Allosterie přejde na jednoduchou kinetiku Michaelise a Mentenové, protože se bude jednat pouze o R stav.