Ruolo dellesperienza e periodi critici nello sviluppo Nel














 richiede l’esperienza")


 possa")






















 si attiva anche durante compiti di")
















 Interburst interval in EEG Reduction in interburst interval, T 1 -T 2")
 Massaged IGF 1 path Massage leads to higher levels of circulating IGF-1")
 P 12 P")





















. Atmospheric 14 C that was released")

























- Slides: 109
Ruolo dell’esperienza e periodi critici nello sviluppo Nel 1693 lo scienziato William Molyneux pose al suo amico, il filosofo Locke, un “jocose problem”.
La domanda di Molyneux era: “immagina un uomo adulto e cieco dalla nascita e che questi abbia imparato a distinguere un cubo da una sfera di avorio (approssimativamente della stessa grandezza) mediante il tatto. Supponi poi che il cieco recuperi la vista, la questione è se servendosi della sola vista, e prima di toccarli, egli saprebbe distinguere la sfera dal cubo”.
Il problema di Molyneux implica una riflessione sul ruolo rispettivamente della specificazione genica e dell’esperienza nell’acquisizione di una funzione e nello sviluppo del comportamento. Problema “nature vs nurture”, la cui domanda di base è: quale è il contributo del patrimonio genetico allo sviluppo di un comportamento e quale è il contributo dell’ambiente e dell’esperienza allo sviluppo di quello stesso comportamento. Il dibattito si è sviluppato fra diverse correnti di pensiero, basti ricordare il dibattito fra Comportamentisti ed Etologi svoltosi intorno alla seconda metà del secolo scorso. Da una parte si sosteneva che ogni comportamento
Lorenz e il concetto di “predisposizione innata ad apprendere”. Lorenz ha sottolineato la fondamentale evidenza che qualsiasi processo di apprendimento deve necessariamente svolgersi entro i limiti ed i vincoli fisiologici di ciascuna specie. Dal momento che le diverse specie presentano differenze morfologiche e fisiologiche e che queste differenze dipendono dalle leggi genetiche della selezione e dell’ereditarietà, la potenzialità di acquisire nuove informazioni attraverso l’apprendimento è a sua volta regolata e programmata in base a vincoli specie-specifici. Un esempio classico di processo di apprendimento geneticamente determinato per avvenire nel momento biologicamente più opportuno è l’imprinting (Horn, 2004).
Oggi noi sappiamo che durante lo sviluppo il ruolo dell’esperienza è fondamentale affinché i circuiti cerebrali, inizialmente formatisi sulla base di specificazioni genetiche, maturino in maniera appropriata a garantire il normale sviluppo delle funzioni cerebrali e del comportamento.
Alla specificazione genica va il ruolo di guidare i processi iniziali dello sviluppo cerebrale e quello della formazione iniziale delle connessioni neurali. All’esperienza specifica dell’individuo ed alle sue interazioni con l’ambiente va il ruolo di guidare le fasi finali dello sviluppo dei circuiti cerebrali, conducendoli allo stato maturo e quindi guidando lo sviluppo delle diverse espressioni del comportamento in maniera individuo-specifica.
Plasticità neurale: capacità dei circuiti nervosi di modificarsi, in maniera funzionale e morfologica, in risposta all’esperienza Cosa cambia nel circuito? L’efficacia delle connessioni sinaptiche si modifica in risposta ai cambiamenti nell’attività elettrica che l’esperienza induce nel circuito Durante lo sviluppo l’esperienza controlla e modifica l’efficacia delle connessioni sinaptiche attraverso l’attività elettrica da essa evocata: se il neurone presinaptico attiva in maniera robusta il suo bersaglio postsinaptico, le sue connessioni si potenzieranno e potranno addirittura aumentare di numero; se invece il neurone presinaptico fallisce ripetutamente nell’attivare il bersaglio postsinaptico, le sue connessioni verranno indebolite o addirittura rimosse.
Durante lo sviluppo l’esperienza controlla e modificala formazione, il mantenimento e l’efficacia delle connessioni sinaptiche in un circuito attraverso l’attività elettrica da essa evocata nel circuito stesso. In questo modo l’esperienza rimodella e raffina le connessioni sinaptiche conducendo alla maturazione dei circuiti. Connettività debole e diffusa Connettività selettiva e robusta sviluppo
Esperienza e sviluppo del cervello: esempi
Da quando il bambino è sensibile all’esperienza?
Il bambino è sensibile all’esperienza già a livello prenatale. Ad esempio, a partire dal terzo trimestre di gravidanza il bambino è in grado di udire stimoli provenienti dall’esterno (evidenze comportamentali, magnetoencefalografia, preferenze precoci). Alla nascita mostra di aver memorizzato gli stimoli uditivi di cui aveva fatto esperienza prenatalmente.
I neonati preferiscono la voce della loro mamma a quella di una donna estranea e la capacità di riconoscere la voce della mamma è acquisita prima della nascita: se viene dato loro da scegliere fra la voce della mamma filtrata in modo da simulare lo spettro di frequenze disponibili in utero e la voce della mamma non filtrata, i neonati preferiscono la prima. I neonati preferiscono anche ascoltare la loro lingua madre rispetto ad una lingua straniera: date le caratteristiche di filtraggio dell’ambiente uterino, è il ritmo e l’intonazione della lingua materna che è stata memorizzata.
Però la percezione è immatura: es. la visione del bambino è molto immatura alla nascita
Sviluppo dell’acuità visiva Acuità visiva normalizzata Uomo Scimmia 1. 0 0. 5 0. 0 0 20 40 60 80 100 0 5 10 15 Ratto Gatto 1. 0 0. 5 0. 0 0 2 4 6 0 Età (mesi) 1 2 Berardi, Pizzorusso, Maffei, 2000
Acuità visiva normalizzata Lo sviluppo dell’acuità visiva (e della visione in generale) richiede l’esperienza Acuità visiva in assenza di esperienza visiva dalla nascita Uomo 0. 5 Sviluppo normale 0. 0 0 20 40 Età (mesi)
Sviluppo delle dimensioni dei campi recettivi nella corteccia visiva Piccolo Adulto
Piccolo Adulto in assenza di esperienza visiva Effetti della mancanza di esperienza visiva sullo sviluppo dei campi recettivi nella corteccia visiva
Durante lo sviluppo la elevata plasticità neurale è cruciale affinché l’esperienza (attività elettrica) possa guidare la maturazione dei circuiti neurali e delle funzioni che da essi dipendono.
I passi che conducono ad una modifica, funzionale e morfologica, duratura dei circuiti neurali in risposta all’esperienza coinvolgono diversi fattori Il recettore NMDA funge da detettore di attività correlata fra il terminale presinaptico ed il terminale postsinaptico Livelli di inibizione Fattori neurotrofici (NGF, BDNF) Protein chinasi (ERK, PKA) Neuromodulatori (Acetilcolina, Noradrenalina) Fattori di trascrizione (CREB) Componenti della matrice extracellulare e proteasi che li degradano Rimodellamento della cromatina conseguente cambiamento nella trascrivibilità di specifici geni Fin qui 19 ottobre 2017 Se uno di questi fattori manca (difetto genetico) l’esperienza può non riuscire a guidare efficacemente la maturazione dei circuiti nervosi
L’esperienza agisce attraverso specifici fattori di plasticità Acuità visiva Esperienza mancante, Sviluppo normale somministrazione di BDNF Normale esperienza, assenza di NGF 1. 0 0. 5 0. 0 età Berardi et al. , (1994)
L’esperienza è particolarmente efficace nel modificare i circuiti nervosi durante periodi critici dello sviluppo
Acuità visiva normalizzata Dopo la fine dei periodi critici l’esperienza non è più in grado di guidare lo sviluppo di una funzione Uomo 1. 0 Acuità visiva a tre anni 0. 5 0. 0 Acuità visiva in assenza di esperienza visiva dai tre anni 0 20 40 60 80 100 420 Età (mesi) Fine et al. , (2003)
Cataratte monoculari neonatali, difetti di refrazione o strabismo non corretti precocemente durante lo sviluppo determinano ambliopia dell’occhio deprivato e forte riduzione della visione binoculare. S =1. 9 9 S = 2. 5 S = 3. 2 S= 4 S=5 S= 6. 3 S=8 S=10 Table at 4 m D= 20 D = 1 6 C = 80 D = 12. 5 D=10 D=8 D = 6. 3 C =3 2 D =5 C = 25 D = 4 C =20 Snellen Fraction is: S/10 or 20/C C=100 C=63 C=50 C=40
Periodo critico per la plasticità della dominanza oculare Effect of MD (for man, recovery from strabismus) Normalized MD effect Human Monkey 1. 0 0. 5 0. 0 1. 0 0 20 40 60 80 100 Cat 0 2 eye opening 0 5 10 15 Rat 1. 0 0. 5 0. 0 0. 5 4 6 0. 0 0 eye opening 1 2 Age (months) Berardi, Pizzorusso, Maffei 2000
Perché la plasticità declina con la chiusura dei periodi critici? Perché la elevata plasticità dei periodi critici ha esaurito il suo scopo, che era quello di consentire all’esperienza di guidare la maturazione dei circuiti neurali
Il declino della plasticità durante lo sviluppo di una funzione correlano visual acuity Monkey 1. 0 0. 5 0. 0 0 20 40 60 80 100 0 5 10 15 Rat Cat 1. 0 0. 5 0. 0 0 2 4 6 0 Age (months) 1 2 Normalized MD effects normalized visual acuity Human Critical period
Man mano che l’esperienza guida la selezione delle connessioni neurali, determinando la maturazione di un circuito e della corrispondente funzione, il circuito e la funzione diventano sempre meno modificabili dall’esperienza. Vantaggi, stabilità Svantaggi, difficoltà di recupero
Gli stimoli: Esperienza A = features B= contour C= relazioni secondo ordine e capacità di riconoscimento dei volti
Differenza tra i valori dei soggetti e la media della popolazione normale
Periodi critici sviluppo ippocampo, amnesia infantile L’ippocampo immaturo risponde all’esperienza di apprendimento attivando fattori di plasticità sinaptica, che consentono la formazione iniziale della traccia di memoria, e soprattutto risponde all’esperienza di apprendimento cambiando l’espressione di questi fattori portandola verso quella trovata nel sistema ippocampale maturo. Tuttavia, bloccando prima dell’apprendimento la funzione di fattori che nel sistema visivo determinano la chiusura dei periodi critici (Huang et al, 1999; Berardi et al. , 2015), si impediva l’effetto antiamnesico della riesposizione ad una componente dell’esperienza iniziale e si impediva anche l’effetto dell’esperienza di apprendimento sulla espressione dei fattori di plasticità sinaptica. Viceversa, aumentando l’espressione dei fattori che determinano la chiusura dei periodi critici visiva in corrispondenza dell’apprendimento promuoveva la formazione di una memoria episodica duratura (Travaglia et al. , 2016). Lavori Cristina Alberini
Le similarità tra questi risultati e quelli trovati nei periodi critici dei sistemi sensoriale consente di proporre che il sistema di memoria dell’ippocampo attraversa un periodo critico simile a quello dei sistemi sensoriali. Durante questo periodo l’ippocampo immaturo è altamente sensibile all’esperienza fornita dall’apprendimento in termini di attivazione dei fattori di plasticità sinaptica, anzi, è più sensibile che ad età successive (Travaglia et al. , 2016 b) ma usa questa sensiblità per maturare. In altre parole, l’esperienza che ha luogo durante questo periodo critico dello sviluppo ippocampale determina cambiamenti nei circuiti ippocampali che servono alla loro maturazione, portandoli ad uno stato di competenza funzionale che li rende finalmente in grado non solo di formare memorie episodiche ma anche di consolidarle a lungo termine e di trasferirle poi alle strutture corticali per il loro mantenimento (Alberini e Travaglia, 2017).
Fig 2. 5
Periodi critici nello sviluppo del sistema serotoninergico per l’emergenza di un fenotipo “ansioso” La presenza e la funzionalità del recettore 1 A per la serotonina (5 -HT 1 A) è centrale al coinvolgimento del sistema serotoninergico nel comportamento ansioso. Recentemente, è stato suggerito che una ipofunzionalità di questo recettore durante un particolare periodo dello sviluppo postnatale possa produrre una alterazione permanente del comportamento ansioso, cosa che una equivalente ipofunzionalità nell’adulto non causerebbe.
E’ possibile correggere un errore di sviluppo una volta chiusi i periodi critici?
Agire sui fattori che “frenano” la plasticità con la chiusura dei periodi critici
Azione farmacologica su un “freno” della plasticità Ch-ABC or P-ase injections 7 days MD Birth P 21 P 23 RS > P 120 (median P 160)
Rimozione farmacologica di un “freno” sulla plasticità corticale nell’adulto: recupero dall’ambliopia 1, 4 Deprived eye Visual Acuity (c/deg) Fellow eye 0, 8 0, 2 * RS-Control RS+ch. ABC Pizzorusso et al. , 2006 I CSPG controllano la plasticità sinaptica strutturale
La modifica delle spine dendritiche rappresenta una traccia permanente di un fenomeno di plasticità La DM durante il periodo critico causa un aumento della densità di spine nei neuroni piramidali dello strato V che rimane anche dopo che il periodo di deprivazione è terminato SB Hofer et al. Nature 000, 1 -5 (2008) doi: 10. 1038/nature 07487
Un secondo episodio di deprivazione monoculare nell’adulto aumenta il volume delle spine che erano state aggiunte dopo la prima deprivazione ma non aumenta ulteriormente il numero delle spine. La cosa più importante è che in questi animali che hanno subito l’episodio giovanile di deprivazione il secondo episodio di deprivazione è efficace nello spostare la dominanza oculare. Normalmente, una deprivazione monoculare è inefficace nell’adulto. L’esperienza durante il periodo critico ha lasciato una traccia a livello sinaptico, per cui l’adulto è suscettibile ad una modifica dell’esperienza che negli altri adulti non ha effetto
Plasticità cross-modale Normalmente i compiti di discriminazione tattile comportano l’uso e la conseguente attivazione dell’area Somatosensoriale. La lettura dell’alfabeto Braille nei ciechi dalla nascita attiva la corteccia visiva primaria.
Nei ciechi dalla nascita la corteccia occipitale (“visiva”) si attiva anche durante compiti di memoria verbale (riprodurre mentalmente parole da una lista memorizzata). I ciechi dalla nascita hanno migliori capacità dei soggetti normali in compiti di memoria verbale: la % di parole di una lista ricordate 6 mesi dopo l’apprendimento è maggiore di quella dei soggetti normovedenti. Questa capacità correla soggetto per soggetto con l’entità di attivazione della corteccia occipitale durante il compito. Amedi et al. , 2002
Ambiente, plasticità e sviluppo del comportamento: il paradigma dell’ambiente arricchito
Cosa si intende per ambiente arricchito La prima definizione formale di ambiente arricchito è stata da Rosenzweig: “una combinazione di stimolazione sociale ed inanimata complessa” (Rosenzweig et al. , 1978).
Ambiente arricchito: condizioni sperimentali § Condizione di impoverimento § Condizione standard di laboratorio (non AA). § Ambiente arricchito con possibilità di compiere esercizio fisico volontario (AA).
Elementi chiave di un ambiente arricchito?
We think that the crucial elements in EE are novelty, cognitive activity, motor activity, social activity, satisfaction and reward
Sviluppo visivo in animali “arricchiti”
Sviluppo precoce dell’acuità visiva in ratti “arricchiti” EE Non-EE normalized VEP acuity 1. 0 0. 8 0. 6 P 19 -20 P 22 -23 P 25 -26 P 28 -29 P 34 -35 P 44 -45 postnatal age (days) Landi et al. , 2007
The first studies employing EE documented an increase in cortical thickness, an increase in spine density and in dendritic tree complexity, as well as effects on neuromodulators such as acetylcholine and noradrenaline and an improvement in learning and memory performance. More recently, a strong EE effect on neural plasticity and in particular on synaptic plasticity has been shown (Sale et al. , 2014)
The steps leading to long term morphological and functional changes in neural circuits in response to experience involve several cellular and molecular factors The first step is the activation of a particular receptor for the neurotransmitter glutamate, the NMDA receptor, which acts as a coincidence detector for the presence of correlated activity between the presynaptic and postsynaptic neuron. Inhibition/excitation balance Neurotrophic factors (NGF, BDNF) Protein kinase (ERK, PKA) Neuromodulators (ACh, De, Ne, 5 -HT) (motivation, attention, interest, pleasure) Transcription factors (CREB) Chromatin remodelling with the ensuing long term changes in gene trascription probability (epigenetic mechanism) Homeostatic plasticity factors Extracellular matrix components and degrading enzymes EE acts on ALL these factors (and also on electrical activity)
Impoverishing the environment reduces IGF 1 in the visual cortex and delays visual acuity development P 12 Visual cortex IGF 1 P 28 visual acuity Impoverished condition Standard Condition IGF-1 cells/mm 2 * VEP acuity (c/deg) 1, 0 * 0, 8 P 18 visual acuity 0, 6 1000 0, 4 SC IMP 0 IMP SC Baroncelli, Narducci, Cenni et al, in preparation
Bidirectional effects of early environmental richness on visual acuity development EE Non-EE normalized VEP acuity 1. 0 IE 0. 8 0. 6 P 19 -20 P 22 -23 P 25 -26 P 28 -29 P 34 -35 postnatal age (days) P 44 -45
EE as “promoter” of the expression of those very same molecules visual experience exploits to drive visual development
L’esposizione ad ambiente arricchito influenza l’espressione di alcuni di questi fattori così precocemente che questi effetti non possono essere attribuiti all’interazione dei cuccioli con l’ambiente e certamente non all’interazione visiva
Enriched pups receive enhanced levels of licking 20 Total licking: mother + helper females % of time 16 12 Standard laboratory condition Enriched environment 8 4 0 1 -2 3 -4 5 -6 7 -8 9 -10 Sale et al. , 2004 Postnatal age (days) This result implies a continuous tactile stimulation provided by adult females on enriched pups
E’ possibile che arricchire l’ambiente in termini di massaggio possa influenzare lo sviluppo visivo?
Massage accelerates brain development and the maturation of visual function Preterm infants Rat pups Circulating IGF-1 levels are increased by massage. Block of circulating IGF-1 (rat pups) blocks massage effects Guzzetta et al. , 2009
Results (infant) Interburst interval in EEG Reduction in interburst interval, T 1 -T 2 12 (sec) * 8 4 0 Massaged Controls Massage Birth 1 wk 1 mo Guzzetta et al. , 2009 3 mo 7 mo 9 mo 1 yr
Results (infant) Massaged IGF 1 path Massage leads to higher levels of circulating IGF-1 and IGFBP 3 IGF-1 0 Controls ** - 40 - 80 IGFBP 3 (% variation) 300 100 -100 Massage Birth 1 wk 1 mo Guzzetta et al. , 2009
Block of IGF-1 blocks massage effects on visual acuity development (rat) P 12 P 25 Massage JB-1 injections P 25 * Visual acuity (c/deg) 0, 8 Visual Acuity (c/deg) * Assessment of visual acuity 0, 7 0, 6 0, 5 0, 4 TS Massage TS+JB 1 Massage Controlli Control + JB 1 Guzzetta et al. , 2009
EE and massage act on recognition memory development At P 18 and P 25, non EE rats remember the explored object for 1 hour, but non for 24 h. P 18 EE and massaged rats remember the explored object for 24 h. Baldini et al. , in preparation
The importance of EE in experience-dependent development and of IGF-1 as a “master mediator” of EE has been recently reinforced by Wang et al. (2013). They have found that a mismatch between two visual developmental processes, ocular dominance development and binocular matching of orientation selectivity development, can be caused by overexpression of BDNF, which acts on ocular dominance development and cortical plasticity decline but is unable to drive binocular matching of orientation selectivity, with negative results for the quality of binocular vision
EE exposure and IGF-1 are able to correct this mismatch, ensuring an harmonic development of all properties of visual cortical neurons. Binocular orientation preference mismatch in BDNF overexpressing mice is corrected by EE or IGF-1
EE, massaggio ed IGF-1 durante il periodo pre-svezzamento determinano anche una riduzione del comportamento ansioso nell’animale adulto (ci torneremo sopra nella lezione prossima).
Questo sottolinea l’importanza dell’ambiente come forza guida nello sviluppo del comportamento e indica che l’ambiente agisce modulando fattori endogeni come IGF-1.
I “neuroni del massaggio” Vrontou et al. , 2013
Ambiente arricchito come promotore di plasticità adattiva: recupero dall’ambliopia?
Non invasive strategies to promote recovery of visual acuity in adult rats: EE EE 15 days RS MD Birth P 21 P 23 > P 120 Electrophysiology, Behaviour Anatomy
EE promotes recovery of visual acuity in adult rats Sale et al. , 2007
Binocular vision, Visual cliff Discrimination index A Exploration test 1. 0 BINOCULAR RS-SC RS-EE 0. 8 0. 6 0. 4 * 0. 2 0. 0 Baroncelli et al. , 2014
Thus, understanding the factors underlying the time course of developmental plasticity and mediating the effects of environment might help in understanding how to harness plasticity towards adaptive outcomes and promote brain repair. Sale et al. , 2014 Fin qui 24 ottobre 2017
La ricchezza dell’ambiente agisce anche su una seconda forma di Plasticità neurale: la neurogenesi ippocampale
Neurogenesi nel sistema nervoso adulto e neuroplasticità La presenza di neurogenesi, ovvero di generazione di nuovi neuroni, nel Sistema Nervoso Centrale adulto è stata portata all’attenzione della comunità scientifica da Fernando Nottebohm nella seconda metà degli anni ottanta. Fino ad allora la neurogenesi era stata considerata come un evento presente solamente durante lo sviluppo del sistema nervoso e, con l’eccezione delle cellule recettoriale olfattive, assente nell’adulto: i neuroni adulti venivano considerati una popolazione non rinnovabile. Non si riteneva quindi possibile che nel sistema nervoso adulto, fatta salva l’eccezione dei neuroni olfattivi, ci fossero cellule capaci di proliferare e dare origine a nuovi neuroni.
Nottebohm aveva invece osservato che nei nuclei del canto degli uccelli che rinnovano ad ogni stagione il repertorio canoro venivano generati nuovi neuroni che andavano quindi a rinnovare la popolazione neuronale di tali nuclei.
In particolare, tali neuroni venivano generati, come durante lo sviluppo del sistema nervoso, in una zona proliferativa (zona ventricolare) e migravano verso la destinazione finale nei nuclei del canto (Alvarez-Buylla e Nottebohm, 1988).
Al fenomeno della neurogenesi si accompagnava, come durante lo sviluppo, quello della morte cellulare programmata (apoptosi), per cui la neurogenesi dell’adulto determinava sostanzialmente il reclutamento di alcuni dei nuovi neuroni nei circuiti neurali dei nuclei del canto in parallelo alla morte di altri neuroni neogenerati o di neuroni generati precedentemente. Il reclutamento di nuovi neuroni coincideva con il periodo dell’anno in cui avveniva l’apprendimento canoro: l’ipotesi proposta fu quindi che la neurogenesi nel cervello degli uccelli adulti fosse al servizio dell’apprendimento vocale.
Successivamente lo stesso autore ha dimostrato l’esistenza di neurogenesi anche nell’ippocampo di uccelli adulti che, in autunno, iniziano a cibarsi principalmente di semi ed a immagazzinarli in numerosi nascondigli ai quali poi tornano, dopo ore o giorni, per recuperarli (Barnea e Nottebohm 1994). La capacità di recuperare il cibo immagazzinato in posti diversi richiede che ciascun uccello abbia una mappa aggiornata del suo territorio e che ricordi dove, al suo interno, ha immagazzinato il cibo. Ancora una volta, l’ipotesi proposta fu che l’aggiunta di nuovi neuroni nei circuiti ippocampali fosse importante per l’acquisizione di nuove memorie, in questo caso memorie di tipo spaziale.
Questa ipotesi è stata proposta anche da Clayton e Krebs in base a studi in un’altra specie di uccelli che immagazzina il cibo;
Questi autori hanno dimostrato che l’esperienza stessa di immagazzinare e dover poi ricordare i siti di immagazzinamento per poter recuperare il cibo determinava un aumento della neurogenesi nell’ippocampo ed una riduzione dell’apoptosi, con il risultato che negli uccelli più esercitati nella memoria spaziale l’ippocampo risultava avere un volume maggiore rispetto al resto del telencefalo (Clayton e Krebs, 1994).
Nella seconda metà degli anni novanta la presenza di neurogenesi è stata dimostrata anche in zone specifiche del cervello di mammiferi adulti, uomo incluso, ed in particolare nel giro dentato dell’ippocampo (vedi Van Praag et al. , 2000). Non c’è invece evidenza chiara della presenza di neuroni neogenerati in zone neocorticali nell’adulto.
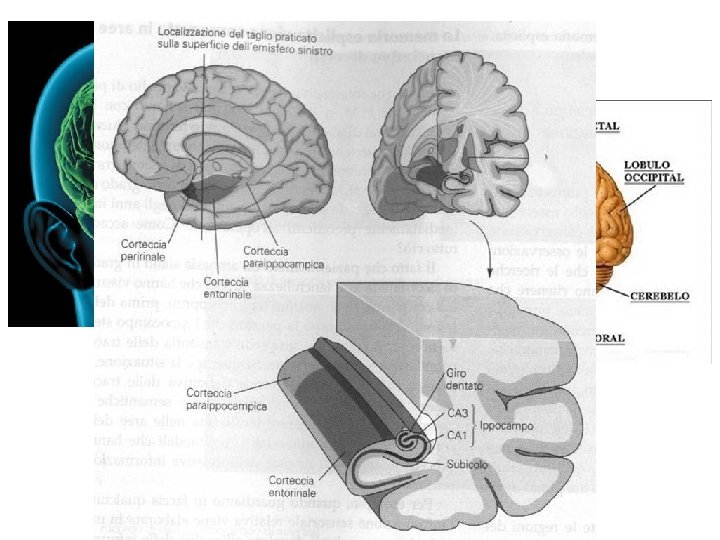
Collaterali di Schaffer CA 1 Fimbria-fornix CA 3 Giro dentato Via delle fibre muscoidi Via perforante Circuito trisinaptico dell’ippocampo. L’informazione dalla corteccia entorinale entra attraverso la via perforante. Le fibre della via perforante formano connessioni sinaptiche eccitatorie sulle cellule dei granuli del giro dentato. Queste ultime danno origine alla via delle fibre muscoidi, che forma connessioni sinaptiche eccitatorie sulle cellule piramidali del campo CA 3. Le cellule piramidali del CA 3 proiettano alle cellule piramidali del campo CA 1 attraverso le collaterali di Schaffer, che formano connessioni eccitatorie. Le cellule del campo CA 1 riproiettano alla corteccia entorinale.
The brain, time-stamped (Spalding et al. , 2013). Atmospheric 14 C that was released during nuclear bomb tests between 1945 and 1963 has been incorporated into the DNA of dividing cells, providing a time-stamp. This has been used to prove adult hippocampal neurogenesis in humans, thereby confirming a particular type of structural and functional brain plasticity involved in higher cognitive function. (News and views, Kemperman et al. , 2013)
By modelling the process of carbon 14 presence in brain cells, the team estimated that we generate around 700 new neurons every day in the DG, roughly the same percentage as in middle aged rodents; most of them die, but some are integrated into DG circuits. The annual turnover rate has been estimated around 1. 75%, which, again, is similar to the levels found in middle-aged rodents. Kheirbeck and Hen, 2013
It has been 56 years since the first evidence of adult hippocampal neurogenesis in the mammalian system was reported and 20 years since it was first demonstrated in the human brain. Despite substantial supporting data, recent contradicting reports by Sorrells and colleagues [2018] and Boldrini and colleagues [2018] show that the very existence of human hippocampal neurogenesis can still be a subject for debate.
Sorrells and colleagues examined 18 adult and 19 perinatal or postnatal samples of postmortem brain tissue obtained from individuals of wide age range (14 gestational weeks to 77 years) with various causes of death (full medical history was not provided by the authors). Upon extensive immunohistochemical analysis of cells labeled with markers for proliferation (Ki-67+), young immature neurons (DCX/PSA-NCAM+), radial glia-like stem cells, and glia in the hippocampus, the authors observed the highest number of proliferating cells and young immature neurons during the first year of life in the dentate gyrus (DG). The authors reported a sharp age-dependent decrease in the number of these cells. Only a few isolated young neurons were observed by 7 and 13 years of age. No young neurons were detected in the DG of adult patients with epilepsy or healthy adults.
By contrast, Boldrini and colleagues examined 28 postmortem hippocampal tissue samples derived from healthy adults ‘without cognitive impairment, neuropsychiatric disease, or (history of medical) treatment’ from 14 to 79 years of age. The authors used similar immunohistochemistry methods as Sorrells and colleagues did to visualize various cell types relevant to hippocampal neurogenesis, including quiescent neural progenitors, proliferating intermediate progenitors, young immature neurons, and mature granule neurons. In contrast to Sorrells and colleagues’ observations, Boldrini and colleagues were able to detect all of the immature and mature cell types in the adult human DG, as shown in previous other studies. Furthermore, they did not observe a substantial decline in the number of these cells with age. The number of cells for each cell type were estimated to be at least thousands per DG subregion, supporting a previous study that showed similar numbers for adultborn hippocampal cells.
At first, it may appear surprising that the two studies used similar methods and reached completely opposite conclusions as to whether adult human hippocampal neurogenesis exists. However, a closer look at the subtle differences in their methods may provide clues to why this might have happened. One of the key strengths of Boldrini and colleagues’ study, not implemented by Sorrells and colleagues, is the use of stereology, which is widely regarded as the gold standard for unbiased quantification in histological studies. The adult brains examined by Sorrells and colleagues also had at least 20 h longer postmortem delay compared with those used by Boldrini and colleagues, which may have further diminished the immunoreactivity of markers such as DCX. .
Furthermore, the majority of control adults studied by Sorrells and colleagues had various diseases, such as cancer and stroke, whereas Boldrini and colleagues took rigorous measures to ensure that their subjects were physically and psychologically healthy. Therefore, it is difficult to say that the findings of Sorrells and colleagues represent adult human hippocampal neurogenesis in the general population, and there is a high chance that Boldrini and colleagues were more likely to have generated accurate estimates of neurogenesis in healthy human adults using a bias-free approach, such as stereology.
However, although Boldrini and colleagues used more reliable methods to study neurogenesis compared with Sorrells and colleagues, both studies share common ground in that they clearly demonstrate the limitations of studying this phenomenon in human postmortem brain tissue. Advancements in non-invasive imaging and biomarker studies would also help us to investigate the role of adultborn neurons at the neural circuitry level and study the functional relevance of these neurons directly in human participants in the context of many interesting subjects that neurogenesis has been previously associated with, such as cognition, mental health, and lifestyle
As two sides of the same coin, the two studies provide a push to the field to develop more advanced tools and models that will generate functionally relevant data for human hippocampal neurogenesis.
Fasi della neurogenesi nell’ippocampo adulto dei roditori
Le cellule che sopravvivono differenziano nel giro dentato, dove si integrano funzionalmente nei circuiti locali ricevendo e stabilendo appropriati contatti sinaptici e durante le prime fasi di questa integrazione sembrano essere particolarmente suscettibili a fenomeni di plasticità sinaptica di tipo LTP (Saxe et al. , 2006; Aimone et al. , 2007). Anche per la neurogenesi presente nell’ippocampo dei mammiferi adulti è stata proposta l’ipotesi che essa sia importante per l’acquisizione di nuove memorie ippocampo dipendenti (van Praag et al. , 2000).
La verifica dell’ipotesi che la neurogenesi ippocampale fosse importante per l’acquisizione di nuove memorie ippocampo dipendenti nei mammiferi ha richiesto molto tempo e molti lavori ed è ancora in corso. I primi studi sono stati di tipo correlativo. In particolare, è stato dimostrato che protocolli di aricchimento ambientale o di solo esercizio fisico, che aumentano la plasticità neurale nell’ippocampo e potenziano le capacità di apprendimento e memoria nei roditori anche anziani, aumentavano la neurogenesi nel’ippocampo (van Praag et al. , 2000). L’effetto di aumento sulla neurogenesi sembrava accompagnato da una riduzione dell’apoptosi nel giro dentato, sia spontanea che indotta da lesione (Young et al. , 1999).
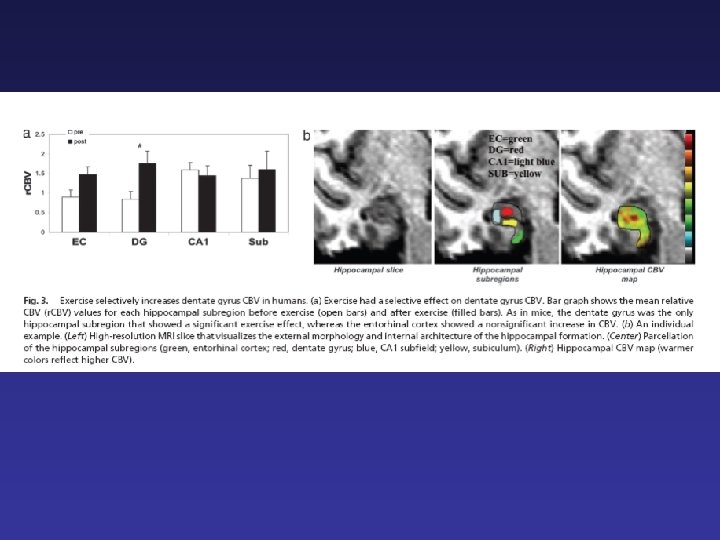
Nel 2007, il gruppo di Gage ha ottenuto interessanti risultati nell’uomo con un protocollo di esercizio fisico (Pereira et al. , 2007). Utilizzando una tecnica di risonanza magnetica che consente di stimare il flusso ematico cerebrale, Pereira et al (2007) hanno mostrato che 12 settimane di training aerobico (circa 1 h al giorno per 4 giorni alla settimana) determinava un aumento del flusso ematico nell’ippocampo e principalmente nel giro dentato
I cambi nel flusso ematico correlavano, soggetto per soggetto, con la fitness cardiopolmonare e con le prestazioni in un compito di memoria ippocampo dipendente.
La stessa tecnica rivela un aumento del flusso ematico localizzato principalmente nel giro dentato di topi liberi di correre sulla ruota e l’aumento di flusso ematico correla con la neurogenesi del giro dentato, valutata anatomicamente.
Gli autori suggeriscono che l’esercizio fisico agisca, a livello ippocampale, principalmente sul flussi ematico del giro dentato, e che la visualizzazione dei cambiamenti del flusso ematico nel giro dentato siano il correlato dell’aumento di neurogenesi indotto dall’esercizio, data la relazione fra angiogenesi e neurogenesi. L’aumento delle prestazioni cognitive dei soggetti che hanno fatto esercizio potrebbe quindi correlare con un aumento di neurogenesi nel giro dentato.
Ruolo della neurogenesi ippocampale nella memoria: 20 anni di ricerca, molte evidenze contrastanti, nessun consenso definitivo.
Gage/Aimone’s hypothesis: temporal cluster of memories New neurons are more excitable and more likely to undergo LTP
Cartoon example of how temporal associations may exist in long-term human memories. The reactivation of an old memory —such as hearing a hit song again years later—can induce the recollection of other memories that were formed at the same time. According to this hypothesis, these memories would have been originally encoded in part by the same set of young neurons, although this recall would most probably be hippocampus independent. Some memories may be general to the time of life—a summer internship, for example. Others may be repeated events that also occurred during that time period, such as visiting a relative. Finally, meaningful personal events may be recalled, such as meeting someone important for the first time. Aimone etal. , 2006
This would be an example of plasticity with a precise role, to create temporal contiguity, which characterizes many of our autobiographical memories. Aimone and Gage, 2009
Neurogenesi ippocampale e pattern separation The current proposal is that hippocampal neurogenesis might subserve specific aspects of memory formation and in particular that DG is responsible for separating memories that are formed in the hippocampus, for encoding temporal context, and for separating and differentiating between overlapping contextual representations (Sale et al. , 2014). Evidenze: van Praag 2012, Gage 2012, Sahay 2011, Hen 2012
Esperienze diverse, cervelli diversi, capacità diverse Evento 2: a casa di mia zia, d’autunno Evento 1: a casa di mia cugina, d’estate Animale non “arricchito”, poche cellule di nuova generazione nell’ippocampo Animale “arricchito”, molte cellule di nuova generazione nell’ippocampo Giudizio: sono ambienti diversi, distinguo nella memoria quando sono andata da zia o cugina Giudizio: è lo stesso ambiente, confondo nella memoria i due eventi Gage 2012
Sahay 2011 Mice with increased adult hippocampal neurogenesis show normal object recognition, spatial learning, contextual fear conditioning and extinction learning but are more efficient in differentiating between overlapping contextual representations, which is indicative of enhanced pattern separation. Ridotta neurogenesi, chiari segni di paura nel contesto 2 Apprendimento a temere il contesto 1 Segni di paura nel contesto 1 Normale neurogenesi, Nessun segno di paura nel contesto 2
Altri ruoli ipotizzati per la neurogenesi ed i suoi cambiamenti plastici dipendenti dall’esperienza (o da fattori genetici) Neurogenesi e consolidamento di sistema
Kitamura 2009 “decreased neurogenesis is accompanied by a prolonged HPCdependent period of associative fear memory. Inversely, enhanced neurogenesis by voluntary exercise sped up the decay rate of HPC dependency of memory, without loss of memory”. Neurogenesi ippocampale ridotta ippocampale aumentata 10 giorni
Altri ruoli ipotizzati per la neurogenesi ed i suoi cambiamenti plastici dipendenti dall’esperienza (o da fattori genetici) Neurogenesi e disturbi d’ansia: Miller ed Hen, 2014: “Early studies found that adult neurogenesis is impaired in models of depression and anxiety and accelerated by antidepressant treatment. This led to theory that depression results from impaired adult neurogenesis and restoration of adult neurogenesis leads to recovery. Follow up studies yielded a complex body of often inconsistent results, and the veracity of this theory is uncertain. “ Affronteremo questo punto nel corso, in relazione al possibile ruolo di differenti livelli di neurogenesi in soggetti con diversi livelli di resilienza.