glyoxylate cycle Oxidative phosphorylation Oxidative phosphorylation Electrotransfer reaction






")
 Oxidative phosphorylation • Electro-transfer reaction in mitochondria • ATP synthesis 氧化磷酸化反應: 1粒線體中的電子傳遞反應")

。 • 2.")



 and reduced (blue) forms. Also")





 + 8 H+")









")






 • The passage of electrons from QH 2")

 • ROS包含超氧歧自由基(Superoxide radical) (.O 2 - )、過氧化氫(hydrogen peroxide) (H")


 • 催化:")












")












")



- Slides: 96
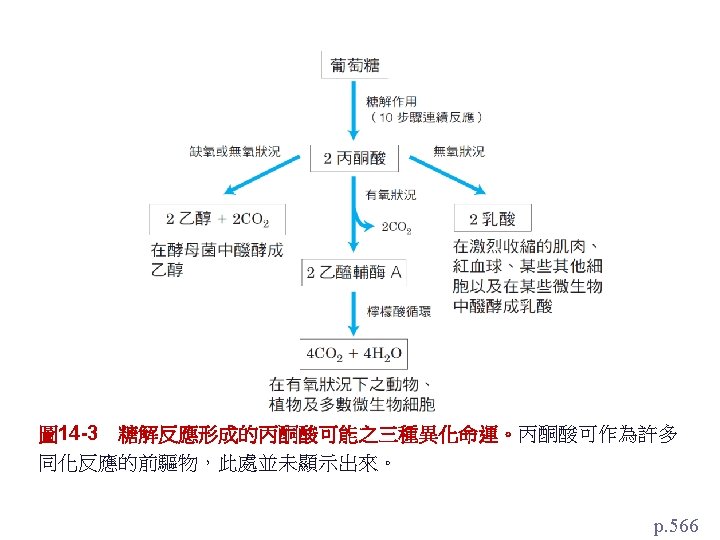
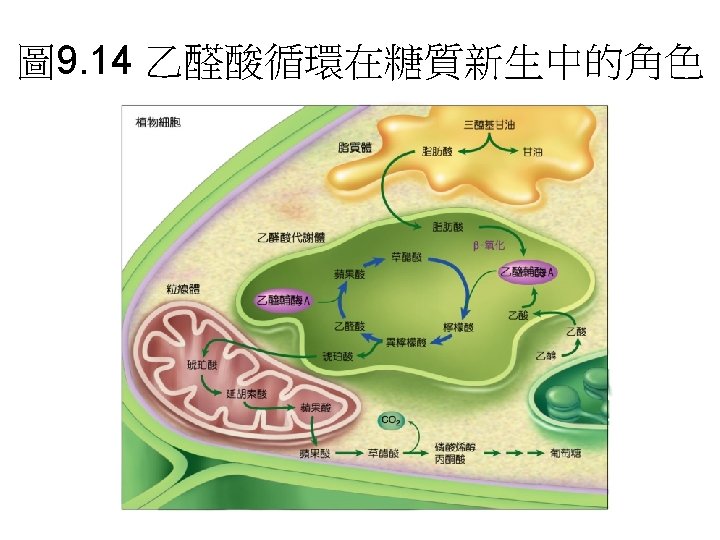
乙醛酸循環(glyoxylate cycle)
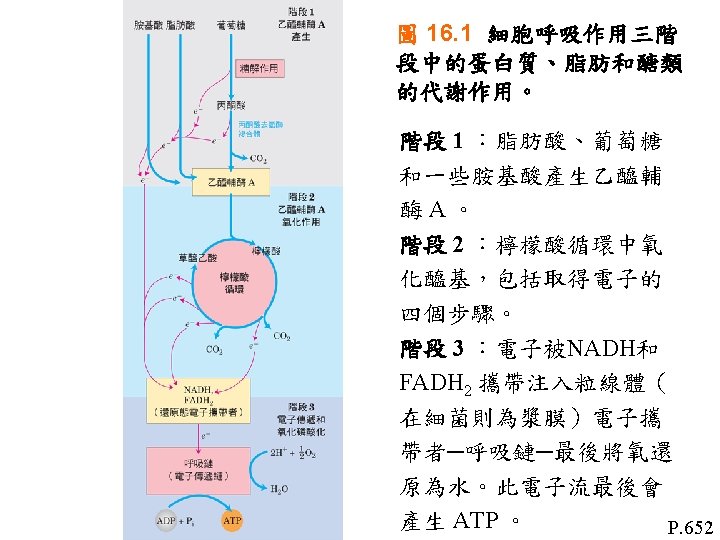
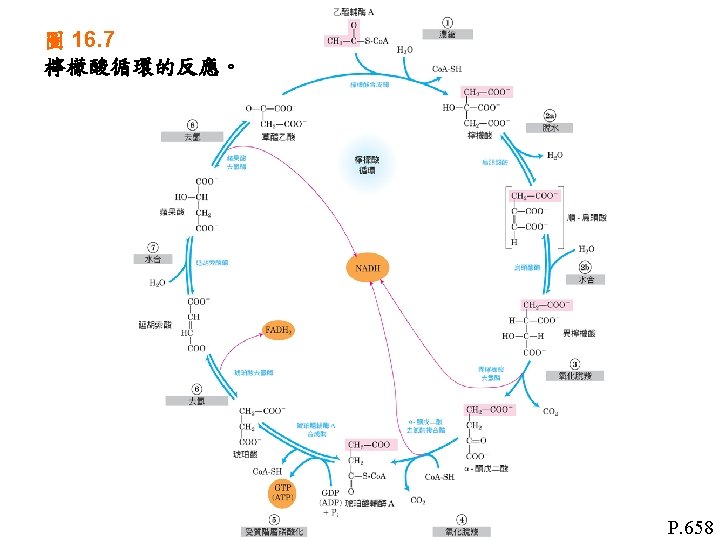
氧化磷酸化反應(Oxidative phosphorylation) Oxidative phosphorylation • Electro-transfer reaction in mitochondria • ATP synthesis 氧化磷酸化反應: 1粒線體中的電子傳遞反應 2 ATP的合成 Regulation of oxidative phosphorylation 氧化磷酸化反應的調節
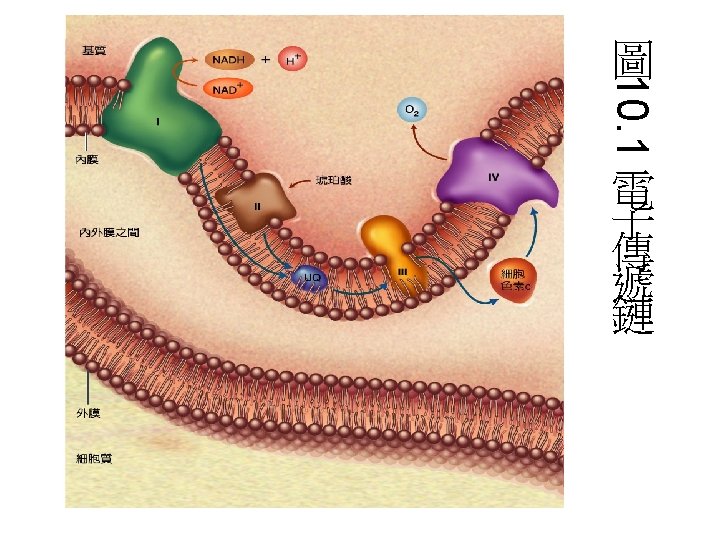
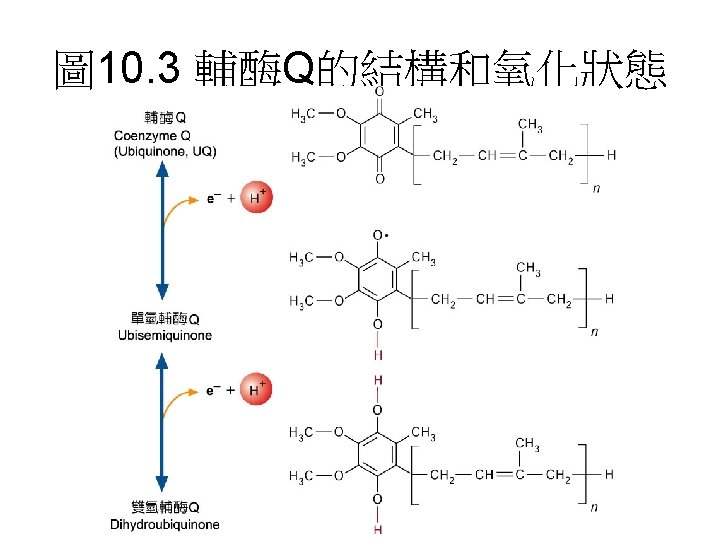
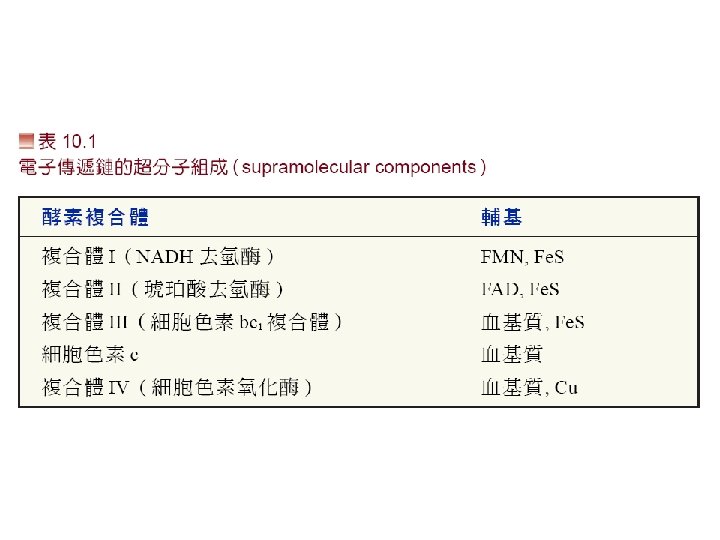
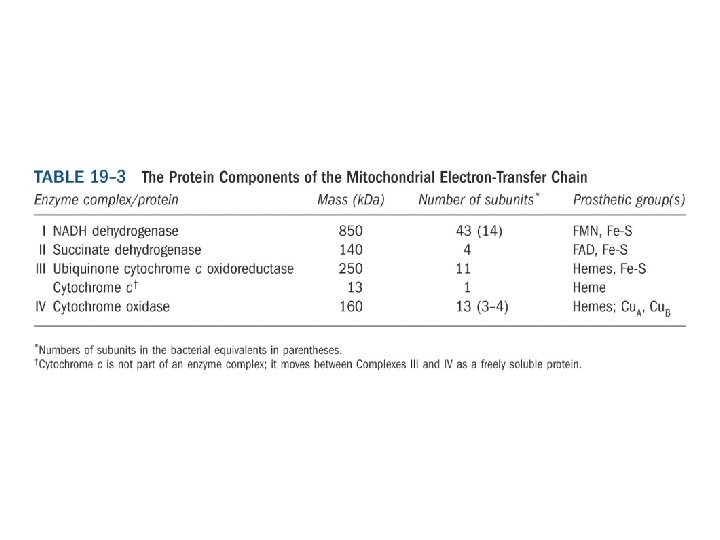
電子傳遞鏈組成分子 • 主要有4個complex組成: • 1. Complex I: NADH dehydrogenase complex 功能: 催化電子由NADH傳至UQ(ubiquinone)。 • 2. Complex II: Succinate dehydrogenase complex: 主要是由Succinate dehydrogenase與2個硫鐵蛋白組 成。功能是催化電子由FADH 2傳至UQ。 • 3 Complex III是由 2個B-type cytochrome、 cytochrome C 1與一個硫鐵分子組成。功能: 將UQH的 電子傳遞至cytochrome C • 4 Complex IV: 由cytochrome oxidase組成 • 功能: 催化 4個電子與O 2形成H 2 O。 • 每個complex皆包含數個蛋白質與prothetic group。 另外還有兩個參與的分子Coenzyme Q與Cytochrome C。
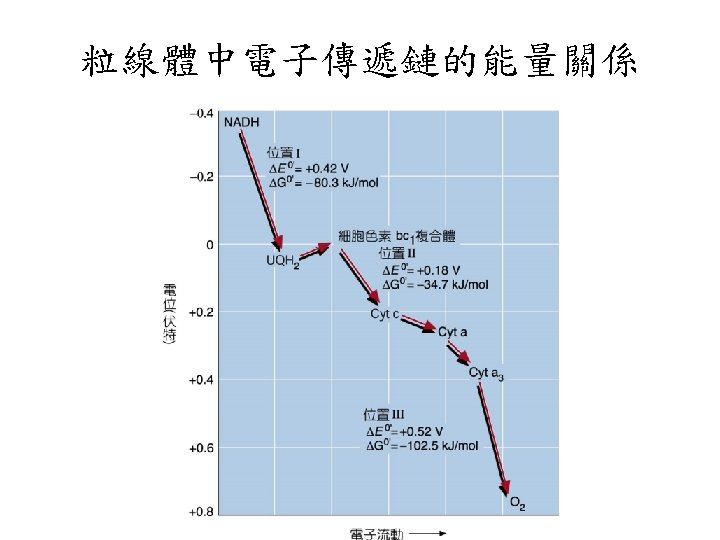
Methods for determining the sequence of electron carriers 還原半電位表
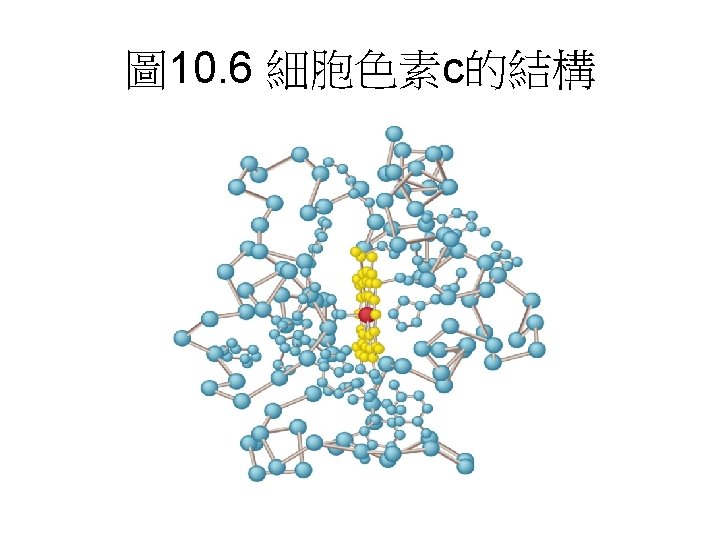
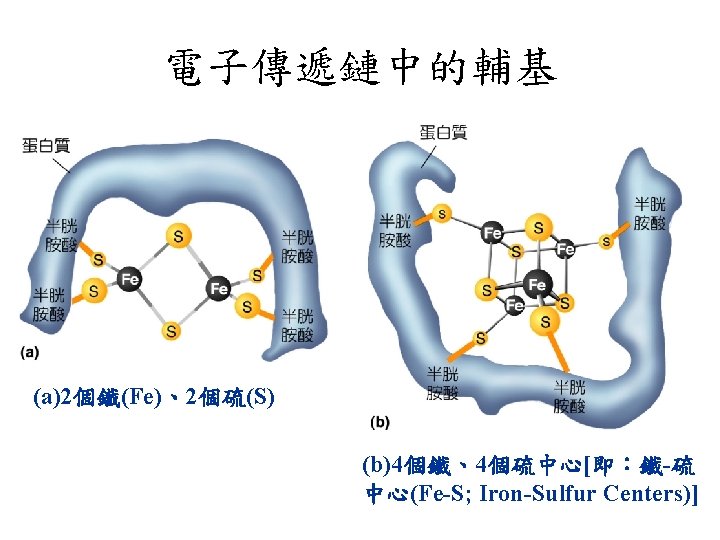
Prosthetic groups of cytochromes. Each consists of four fivemembered, nitrogen-containing rings in a cyclic structure called a porphyrin. The four nitrogen atoms are coordinated with a central Fe ion, either Fe 2+ or Fe 3+.
Absorption spectra of cytochrome c in its oxidized (red) and reduced (blue) forms. Also labeled are the characteristic a, b, and g bands of the reduced form.
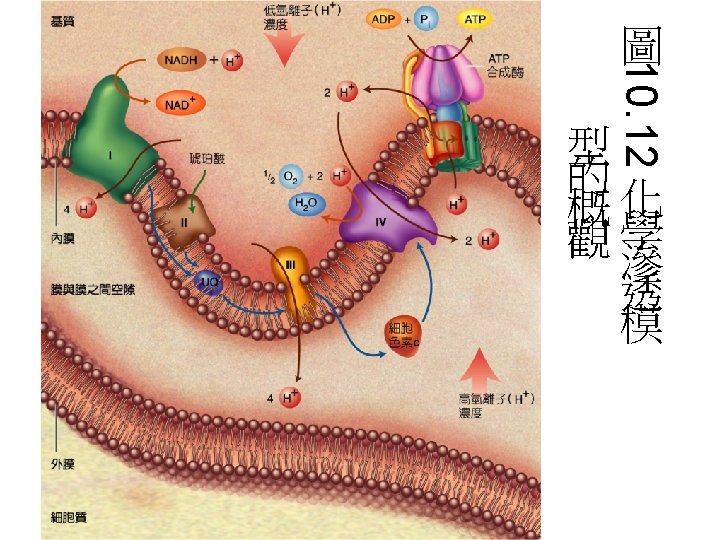
Summary of the flow of electrons and protons through the four complexes of the respiratory chain The vectorial equation for the process is: NADH + 11 HN+ + 0. 5 O 2 → NAD+ + 10 H+P + H 2 O
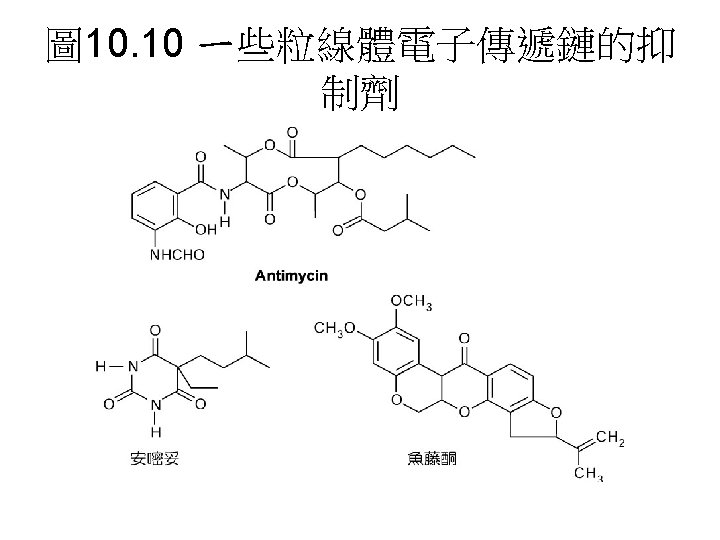
In the presence of an electron donor and O 2, each inhibitor causes a characteristic pattern of oxidized/reduced carriers; those before the block become reduced (blue), and those after the block become oxidized (red). 電子傳遞鏈 中電子傳遞順序與其阻斷劑。
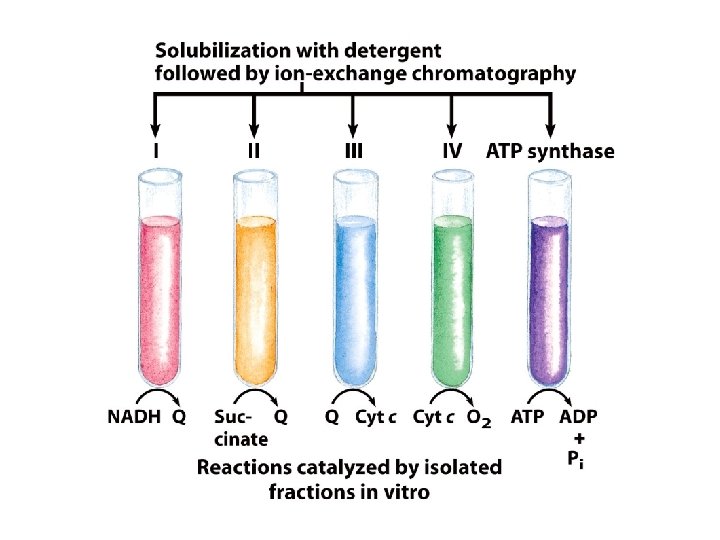
Electron carriers function in multienzyme complexes The electron carriers of the respiratory chain are organized into membrane-embedded supramolecular complexes that can be physically separated. 呼吸鏈的電子攜帶者是位於膜中的巨大蛋白 複合物,且可分離出。
Separation of functional complexes of the respiratory chain 分離呼吸鏈具功能的複合體
The overall reaction catalyzed by Complex IV 4 Cyt c (reduced) + 8 H+ + O 2 → 4 cyt c (oxidized) + 4 H+ + 2 H 2 O Complex IV含銅離子,亦具兩種作用: 1電子 傳遞。2質子的輸送。
Summary of the flow of electrons and protons through the four complexes of the respiratory chain The vectorial equation for the process is: NADH + 11 HN+ + 0. 5 O 2 → NAD+ + 10 H+P + H 2 O
Proton-motive force
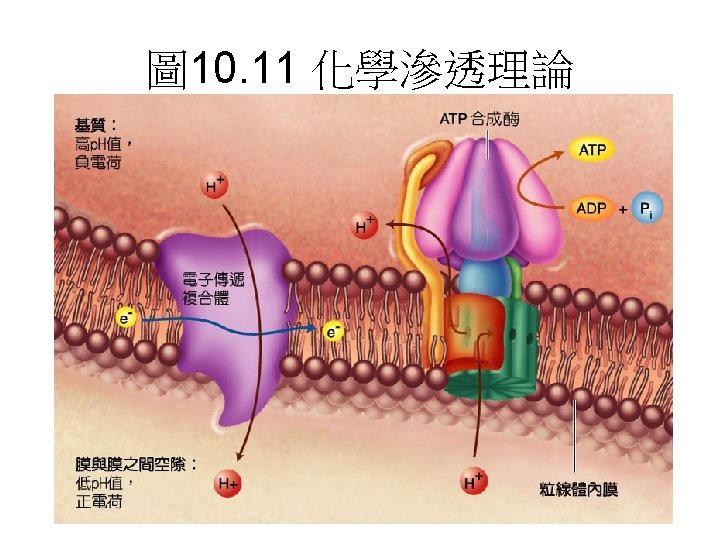
Mitchell假設電子的傳遞導致自由能的釋 致,由ETC產生的質子趨動能會伴隨ADP與 Pi合成ATP。 Because the transfer of two electrons from NADH to O 2 is accompanied by the outward pumping of 10 H+, roughly 200 k. J of 220 k. J released by oxidation of a mole of NADH is conserved in the proton gradient. 估算 1 mole NADH約可打出 10 mole個質子, 花費約200 -220 kj 的能量。
Chemiosomitic model
圖 10. 14 ATP合 成
Two chemical uncouplers of oxidative phosphorylation
圖 10. 13 去偶合劑(uncoupler)
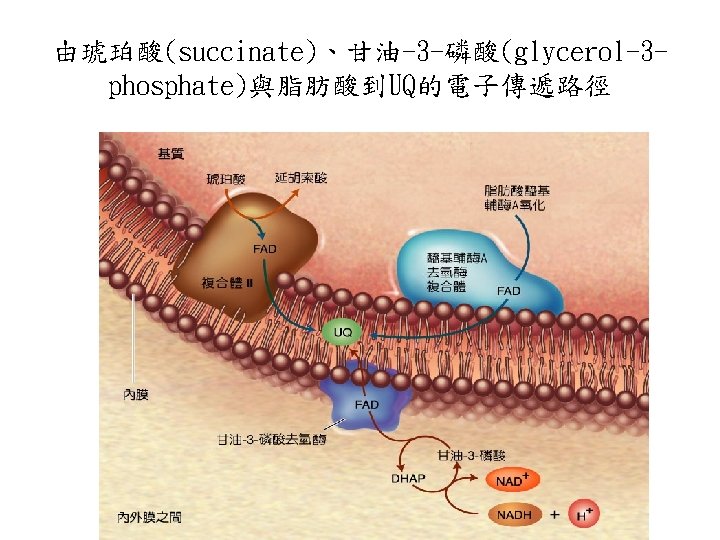
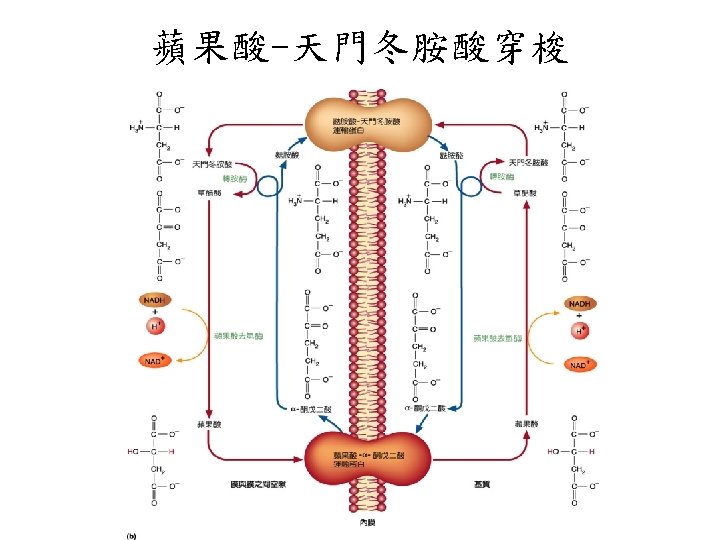
穿梭機制從細胞質的NADH轉移電子到呼 吸鏈: Glycerol phosphate shuttle
Glycerol 3 -phosphate shuttle
Malate-asparate shuttle
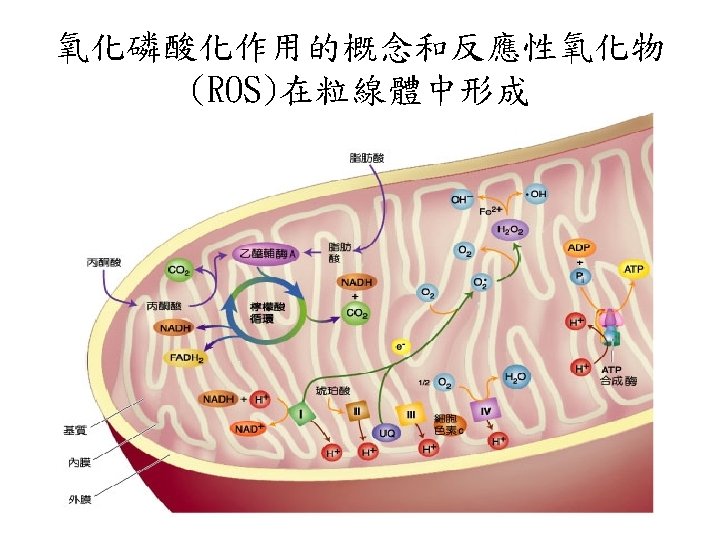
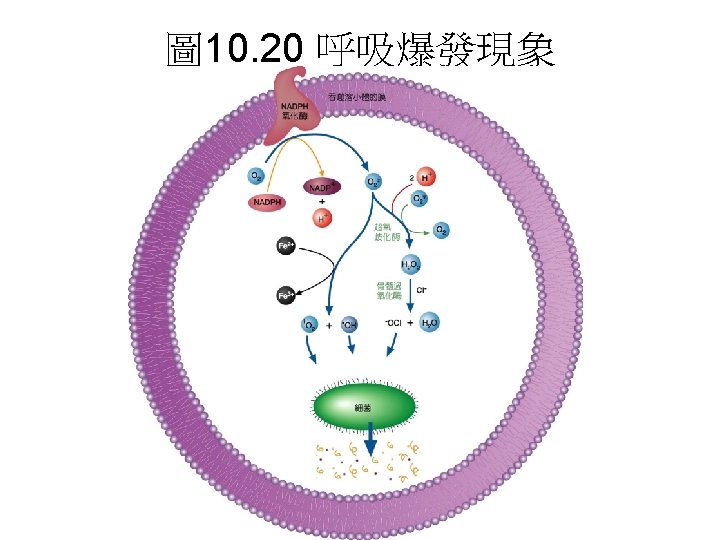
Mitochondrial production and disposal of superoxide(超氧歧的生成與清除) • The passage of electrons from QH 2 to cytochrome b. L through Complex III, and passage of electrons from Complex I to QH 2, involve 醌自 由基(the radical Q) as intermediate. The the radical Q can, with a low probability, pass an electron to O 2 in the reaction O 2 + e- →. O 2 粒線體會產生superoxide: 在複合體I電子傳遞的過程 中,會產生醌自由基再轉移給氧形成超氧歧 (superoxide ·O 2 - )
• The superoxide free radical generated, ·O 2 - is very reactive and can damage enzymes, membrane lipids, and nucleic acids. • Superoxide含自由基對酵素、膜與核酸具 破壞性。
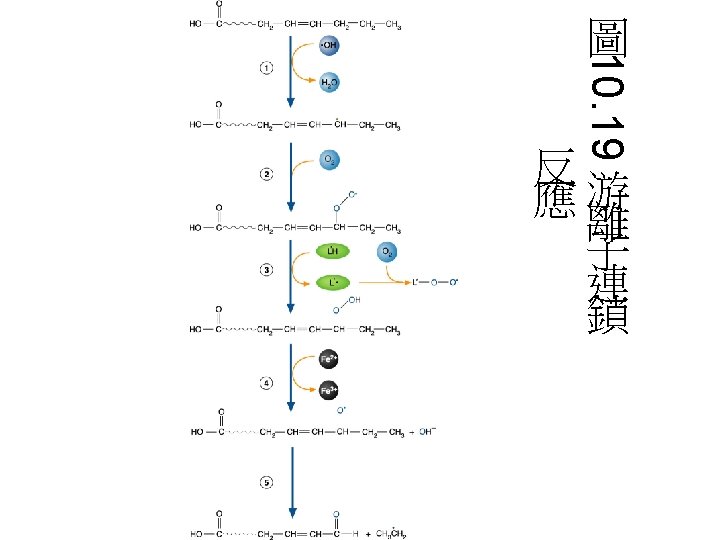
活氧物質: Reactive oxygen species (ROS) • ROS包含超氧歧自由基(Superoxide radical) (.O 2 - )、過氧化氫(hydrogen peroxide) (H 2 O 2)、氫氧自由基(hydroxyl radical) (.OH)與單一氧原子(singlet oxygen) (1O 2)。 • 因為ROS相當reactive,易引起一些鏈鎖反 應。如lipid peroxidation reaction: hydroxyl radical會攻擊一些脂肪酸,引 起鏈鎖反應。食品加 時常會促進這種 radical chain reaction,常需加入抗氧 化劑,阻止radical chain reaction的反應。
阻止氧化壓力的機制 • 一些酵素具有阻止oxidative stress的功 能,如superoxide dimutase、catalase與 glutathione peroxidase。 • SOD (superoxide dimutase) • 催化: 2 O 2 + 2 H+ H 2 O 2 + O 2 • 在入類中,Cu-Zn SOD 位於細胞質內 • Mn SOD則在粒線體內,兩者是同功酶
• Catalase: 是一個含heme的酵素 • 催化: 2 H 2 O 2 2 H 2 O + O 2 , RH 2 + H 2 O 2 R + 2 H 2 O • Glutathione peroxidase: 是一個含Selenium 的酵素,也是控制細胞過氧化物濃度的重 要酵素。 • 催化: 2 GSH + ROOH GSSG + ROH + H 2 O • 2 H 2 O 2 2 H 2 O + O 2
• Glutathione reductase再將GSSG還原成 GSH • GSSG + NADPH + H+ 2 GSH + NADP+ • NADPH主要是由pentose phosphate pathway提供
Mitochondrial production and disposal of superoxide
ATP synthase • Mitochondrial F 1 has nine subunits of five different types, with the composition a 3 b 3 gde. • Each of the three b subunits has one catalytic site for ATP synthesis. • The crystallographic determination of the F 1 structure by John E. Walker and colleagues revealed structural details very helpful in explaining the catalytic mechanism of the enzyme. ATP synthase 的結晶結構由John E. Walker 等解出。
Diagram of the Fo. F 1 complex, deduced from biochemical and crystallographic studies
Binding-change model for ATP synthase
Chemiosmotic coupling allows nonintegral stoichiometries of O 2 consumption and ATP synthesis • If 10 protons are pumped out per NADH and 4 must flow in to produce 1 ATP, the proton-based P/O ratio is 2. 5 for NADH as the electron donor and 1. 5 for succinate. • 1 NADH可產生 2. 5 ATP • 1 FADH 2可產生 1. 5 ATP
Adenine nucleotide and phosphate translocases (located in the inner mitochondrial membrane)
Uncoupled mitochondria in brown fat produce heat • Most newborn mammals, including humans, have a type of adipose tissue called brown fat in which fuel oxidation serves not to produce ATP but to generate heat to keep the newborn warm. • 大部分哺乳動物的幼兒,其脂肪組織具 有棕色脂肪。可以行脂肪氧化,非產生 ATP而產熱。因為棕色脂肪粒線體膜上具 有獨特的蛋白Thermogenin又稱 uncoupleing protein,會破壞質子梯度。
Mitochondrial P-450 oxygenases catalyze steroid hydroxylations • Mitochondria are the site of biosynthetic reaction that produce steroid hormones, including the sex hormones, glucocorticoids, mineralocorticoids, and vitamin D hormone. • 粒線體是合成類固醇與維生素D的胞器。 • These hormones are synthesized from cholesterol or a related sterol in a series of hydroxylations catalyzed by the enzymes of the cytochrome P-450 family. 這些荷爾 蒙是由膽固醇經一系的羥化反應而成,而 催化這些反應的酵素為cytochrome P-450 家族的成員。
• In the hydroxylation reactions, one atom of molecular oxygen is incorporated into the substrate and the second is reduced to H 2 O: R-H + O 2 + NADPH → R-OH + H 2 O + NADP+ • 羥化反應的方程式如上。
Mitochondrial P-450 oxygenases catalyze steroid hydroxylations • Cytochrome P-450 enzymes are situated in the inner mitochondrial membrane with their catalytic site exposed to the matrix. • All of P-450 enzyme have a critical heme group. Its absorption at 450 nm gives this family its name。 • Cytochrome P-450酵素位於粒線體內膜,其催 化的位置曝露於基質中。具有血質(heme), 因對 450奈米具有強的吸收故名之。
• The path of electron flow in the mitochondrial P-450 system is involved a flavoprotein and an iron-sulfur protein that carry electrons from NADPH to the P-450 heme. P-450系統的電子流動路徑包含黃素蛋白、 硫鐵蛋白與NADPH。
Mitochondria of adrenal gland, specialized for steroid synthesis
Another large family of P-450 enzymes • Found in the ER of hepatocytes. 另一大族群則 位內質網,催化反應類似於粒線體的P-450 enzymes。主要用於解毒代謝。 • Catalyze reactions similar to the mitochondrial P-450 reactions. • Their substrates include a wide variety of hydrophobic compounds, many of which are xenobiotics-compounds not found in nature but synthesized industrially.
• Hydroxylation of the hydrophobic compounds makes them more water soluble, and they can then be cleared by the kidneys and excreted in urine. • 羥化不溶於水的疏水性的化合物,可增加 其溶解度,有利於排出體外。
Path of electron flow in mitochondrial cytochrome P-450 reactions in adrenal gland
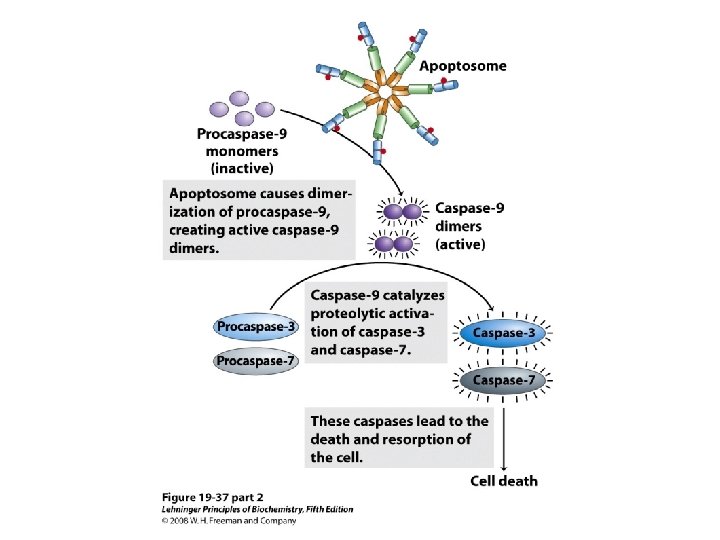
The role of mitochondria in apoptosis • Besides their central role in ATP synthesis, mitochondria also participate in processes associated with cellular damage and death. 除了 合ATP的角色,粒線體也參與細胞損壞與死亡的 過程。 • Apoptosis: Programmed cell death, in which a cell brings about its own death and lysis, signaled from outside or programmed in its genes, systematically degrading its own macromolecules. 細胞凋亡,是計劃性的細死亡。 當細胞接受到死亡訊息,會增加粒線體外膜的通 透性。有利 於cytochrome c 漏出,漏出的 cytochrome c會活化caspase 9,導致細胞凋亡的 啟動。
• When a cell receives a signal for apoptosis, one consequence is an increase in the permeability of the outer mitochondrial membrane ( the opening of the permeability transition pore complex (PTPC)), allowing escape of the cytochrome c normally confined in the intermembrane space. • The released cytochrome c activates one of the proteolytic enzymes (caspase 9) responsible for protein degradation during apoptosis.
Role of cytochrome c in apoptosis
Mitochondrial genes • Mitochondria contain their own genome, a circular double-stranded DNA (mt. DNA) molecule. 粒線體內有一環狀的 雙股螺旋的DNA。共 16569 bases,無 intron且內含 37個基因。隨細胞分裂而分 裂。只有13 genes被轉譯。絕大部分 (95%以上)的粒線體蛋白是由染色體中 的基因所轉譯再送至粒線體中。
Mitochondrial genes and mutations
Heteroplasmy in mitochondrial genomes