Analisi quantitativa delle sequenze di DNA Vincenzo Nigro






 è dominante se l’eterozigote")











 1: 50. 000 nati • Pianto acuto e")





 \"addormentato\", il più")






")









- Slides: 55
Analisi quantitativa delle sequenze di DNA Vincenzo Nigro Dipartimento di Patologia Generale Seconda Università degli Studi di Napoli Telethon Institute of Genetics and Medicine (TIGEM)
Genome Variation SSR Short Sequence Repeats ACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATAT ATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGA CACACACACACACA GACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCT CCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCG AGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCAC ACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTC Allele 1 ACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATAT ATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGA CACACACACACACACA GACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCT CCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGAC ACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGC Allele 2 ACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATAT ATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGA CACACACA GACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCT CCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATAT AGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTG ACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATA Allele 3
ACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTC CGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGC GACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTA GCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCG CACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCT CTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACCGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGAT ATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGAACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGC TCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGC GCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGG CTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCG ATATAGCTCGCGACACAGA TATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCAC ACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATAT ATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGA GACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACA CCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATA TAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGT AGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGA CGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAG CGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGAC GTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAG ACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCC CTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACACACAGATATATAGCG CTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGATAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCT AGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCT AGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTC GCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACC GCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACC GCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTC GAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGA AACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACACACAGATATATAGCGCTCCCTGA AACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGAC ACACACAGATATTATAGCTCGCGACACAGATATATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAG ACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATA GCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACA CGTGCTAGCTCCTCTCGAGACGTTATAGCTCGCGACACAGATATATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTA GCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGAC ACACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCG AGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGC ACACCGCTCGAGATAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCT CGATATAGCTCGCGACACAGATATATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGC TCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCT GAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACAC GTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTC ACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACAC AGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCT CTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGA
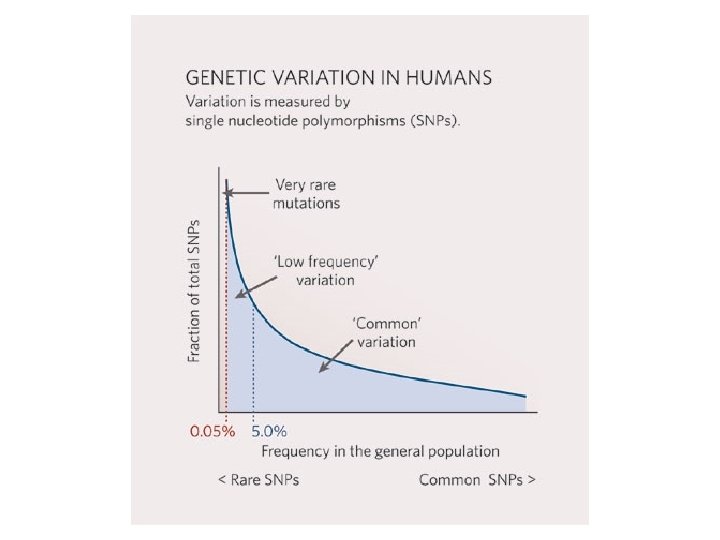
ACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTC CGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGC GACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTA GCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCG CACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCT CTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACCGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGAT ATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGAACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGC TCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGC GCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGG CTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCG ATATAGCTCGCGACACAGA TATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCAC ACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATAT ATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGA GACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACA CCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATA TAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGT AGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGA CGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAG CGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACTATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACCGAGACGTAGGGCTCTCGATATA GCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGC TAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCG ACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAG CTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCT AGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACA GCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACA GCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGAC GTAGGGCTCTCGATATAGCTCGCGACA CACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTC CTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACA GATATATAGCGGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGATAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCT CCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACACACAGATATATAGCGCT CCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGA CCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCT CGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGTAGGGCTCTCGATATAGCT CGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTC CGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCT CGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATA TATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAG ATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACAC CGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACACACAGATATATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACAC AGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATAT AGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTTATAGCTCGCGACACAGATATATAGCGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTG AAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGACGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTGACACGTGCTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGACACAGATATATAGCGCTCCCTGAAACAGCTCCGACACAGCTCGCACACCGCTCGAGACCTTAGCTCCTCTCGAGACGTAGGGCTCTCGATATAGCTCGCGA Copy Number Variation 10% of the human genome could vary in copy number 1 2
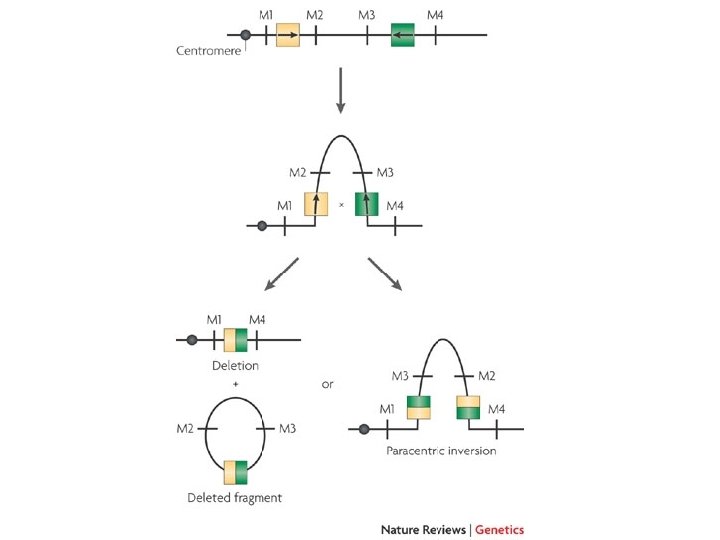
grandi delezioni
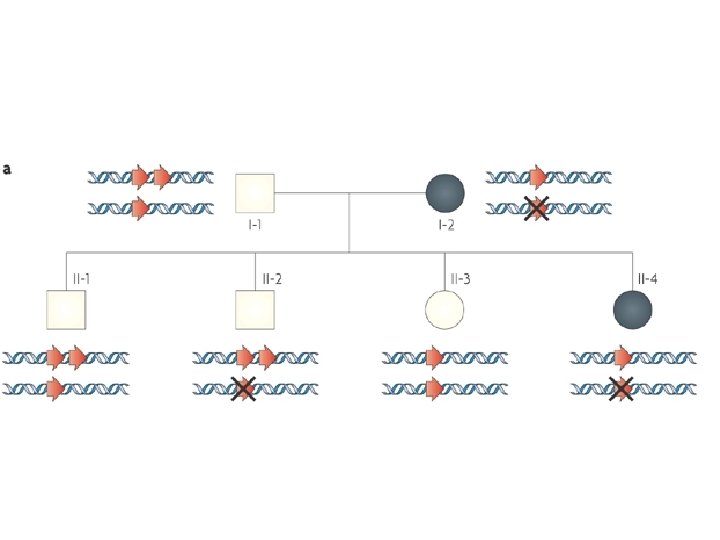
Ipotesi sulla variabilità del fenotipo di 6 individui diversi in caso di trisomia 21 in presenza di varianti alleliche fenotipo Livello di espressione
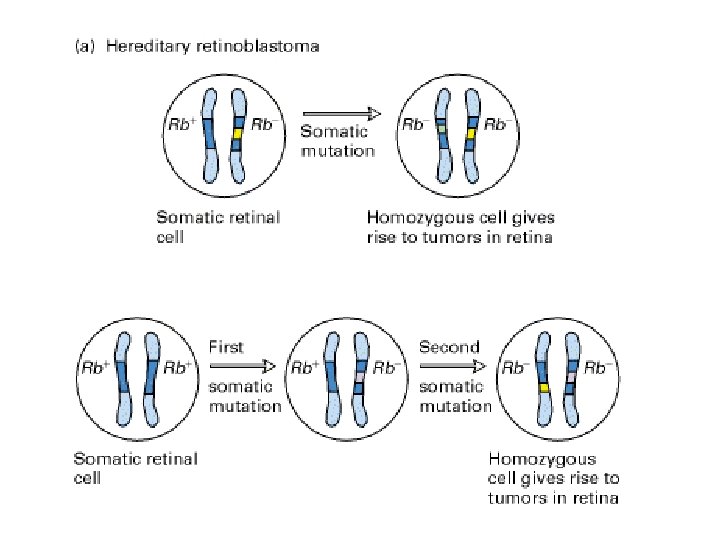
dominanza e recessività • in genetica, il carattere (o l’allele) è dominante se l’eterozigote è indistinguibile dall’omozigote • in medicina la malattia è: – dominante: fenotipo clinicamente manifesto con 1 allele mutato – recessiva: fenotipo clinicamente manifesto con 2 alleli mutati (omozigote o eterozigote composto)
5 effetti di un allele amorfo e ipomorfo causati dalle delezioni • nullo o amorfo = nessun prodotto genico • ipomorfo = ridotta quantità/attività • ipermorfo = aumentata quantità/attività • neomorfo = nuova quantità/attività • antimorfo = quantità/attività antagonistica (dominante negativo)
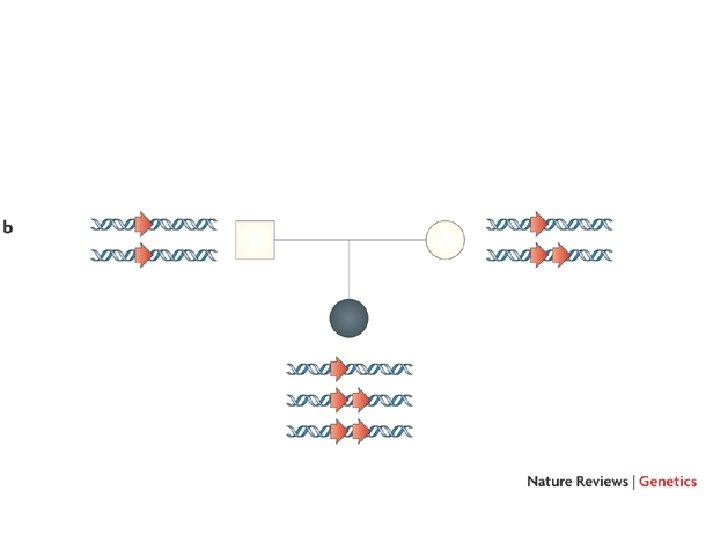
Nella condizione A un fenotipo si osserva solo se entrambi gli alleli sono colpiti (omozigosi o eterozigosi composta) Nella condizione B si osserva un fenotipo già in eterozigosi per aploinsufficienza
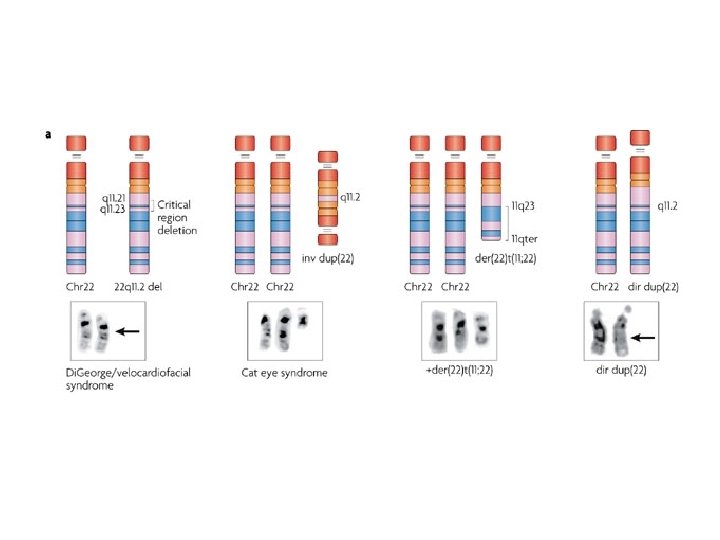
In caso di delezioni del cromosoma X nei maschi si osserva direttamente in fenotipo come sindrome da geni contigui In caso di delezioni autosomiche in eterozigosi molto spesso il dosaggio dimezzato non è causa di malattia. Quando si osserva una sindrome da delezione, è risolutivo trovare la stessa sindrome causata da una mutazione puntiforme in uno solo dei geni. Se questo non c’è mai, la sindrome esiste solo come somma di più difetti
Malattie autosomiche dominanti causate da delezioni basi biochimiche delle delezioni dominanti 1. il livello dimezzato di prodotto genico è insufficiente a mantenere il fenotipo 2. il difetto eterozigote diviene omozigote a livello delle cellule dei tessuti periferici (LOH) 3. Un solo allele è espresso per imprinting dell’altro
aploinsufficienza • insufficiente quantità di prodotto genico causata da una mutazione in eterozigosi • la mutazione è di tipo allele amorfo o ipomorfo • colpisce geni per i quali il 50% di prodotto genico non è abbastanza per garantirne la funzione • spesso un dosaggio preciso è richiesto ai fattori di trascrizione e alle molecole di segnale espressi nel corso dello sviluppo
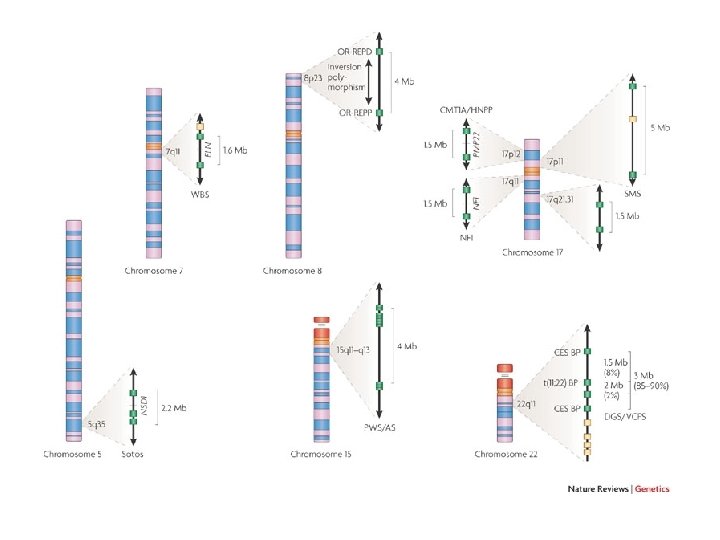
Williams-Beuren • prevalenza alla nascita 1/75001/20. 000, ma può non essere diagnosticata
Williams una delezione tipica
Williams genetica • delezione “de novo” • trasmissione autosomica dominante • delezione di 1. 6 MB da 21 geni contigui in eterozigosi a 7 q 11. 23 – gene dell’elastina – LIM kinase 1 (LIMK 1) – CLIP-115 che lega i microtubuli – Fattori di trascrizione GTF 2 IRD 1 – effetto posizionale su altri geni circostanti la delezione
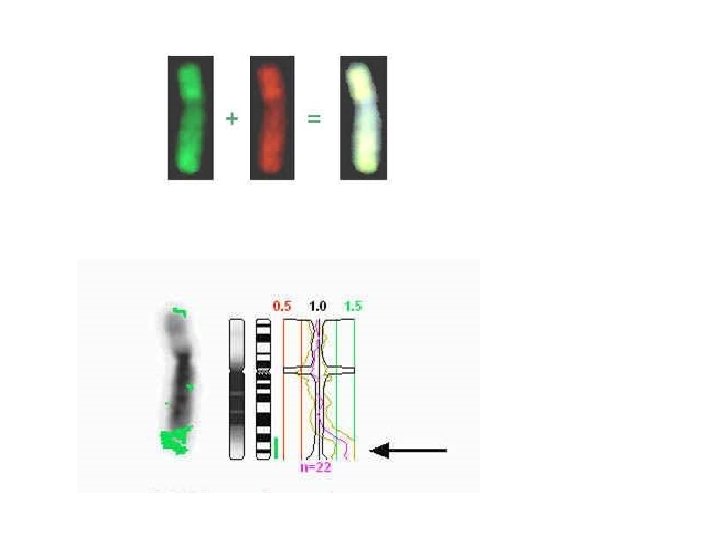
Williams FISH delezione 7 q 11. 23 nrilevabile mediante FISH ma non cariotipo
Wolf-Hirschhorn genetica • delezione “de novo” di circa 4 MB • le delezioni sono più frequenti nella linea germinale maschile • trasmissione autosomica dominante • Regione critica di 165 kb di molti geni contigui in eterozigosi a 4 p 16. 3
Wolf-Hirschhorn • • Scarso accrescimento Ritardo mentale, ipotonia Labbro leporino Conformazione ad elmo di guerriero greco
Sindrome 5 p- (cri du chat) 1: 50. 000 nati • Pianto acuto e flebile • Caratteristiche principali: – Ritardo di crescita – Microcefalia ed ipertelorismo – Ipotonia, diastasi dei retti – Deficit intellettivo e del linguaggio
Sindrome 18 q- • • ipotonia microcefalia ipoplasia della regione centrale del volto ritardo mentale e sordità
Imprinting
Imprinting • Nelle cellule germinali primordiali l’imprinting viene cancellato del tutto e il DNA è demetilato • Successivamente nella linea germinale maschile si determina un pattern di imprinting che in alcuni loci è complementare a quello della linea germinale femminile • I cromosomi su cui avviene l’imprinting (7, 11, 15) manterranno questo pattern e lo riprodurranno ad ogni mitosi • Si potranno sempre distinguere l’espressione genica del cromosoma materno e paterno
Disomia uniparentale • Due copie dello stesso cromosoma sono ereditate dallo stesso genitore • Spesso questo avviene attraverso un fenomeno transitorio di trisomia, seguito dalla perdita del cromosoma singolo e mantenimento del cromosoma doppio
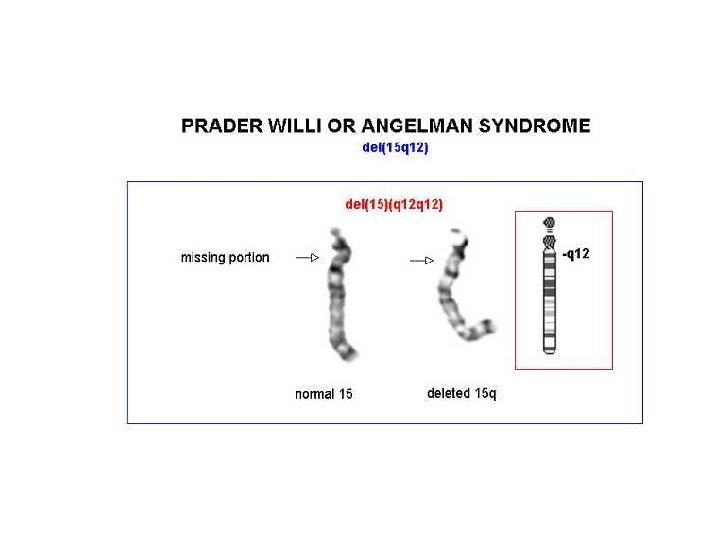
Angelman • 70% dei casi delezione della regione cromosomica 15 q 11 -q 13, che è soggetta al fenomeno dell'imprinting del cromosoma paterno • Il gene materno (l'unico espresso) può essere alterato con 4 meccanismi noti: – – delezione disomia uniparentale paterna difetti nell'imprinting mutazioni a carico del gene UBE 3 A (ubiquitin ligasi) • La diagnosi è clinica e il difetto genetico non si identifica nel 20% dei casi
Angelman • "happy puppet syndrome" si può identificare in Cucciolo (Dopey) "addormentato", il più giovane dei nani che non ha mai imparato a parlare • ritardo mentale con assenza del linguaggio, difficoltà nell'equilibrio, eccessivo buon umore
Angelman • L'incidenza è 1/20. 000 nati • crisi epilettiche e comunque alterazioni dell'EEG e microcefalia relativa
Prader-Willi • • • iperfagia>obesità eccessiva assunzione di liquidi reazioni abnormi ai sedativi acromicria, criptorchidismo insensibilità al dolore, lesioni cutanee sbalzi di umore
Prader-Willi 1/15. 000
Distrofia muscolare Duchenne/Becker DMD Duchenne - 1/3, 500 maschi • Insorgenza -- Infanzia - tra 2 e 6 anni • Sintomi – Debolezza generalizzata e danno muscolare prima agli arti e al tronco, polpacci ingrossati • Progressione – Lenta ma inesorabile. Colpisce tutti i muscoli volontari. Sopravvivenza fino a 25 -30 anni BMD Becker - 1/10, 000 maschi • Insorgenza – Adolescenza o dopo • Sintomi – Identici alla DMD ma più attenuati. Vi è coinvolgimento cardiaco significativo • Progressione – Più lenta e più variabile della distrofia di Duchenne con buona aspettativa di vita
Le delezioni intrageniche del gene della distrofina mandano fuori cornice la lettura delle triplette quando gli esoni cancellati contenevano un numero di nucleotidi che non è multiplo esatto di tre (1, 2, 4, 5, 7, 8, 10, 11 ecc). Questo causa la distrofia di Duchenne
Le delezioni intrageniche non alterano la cornice di lettura portano alla distrofia muscolare di Becker o ad un apparente buona salute. Forniscono informazioni per preparare delle microdistrofine per la terapia genica
Perdita di eterozigosità LOH (loss of heterozygosity)
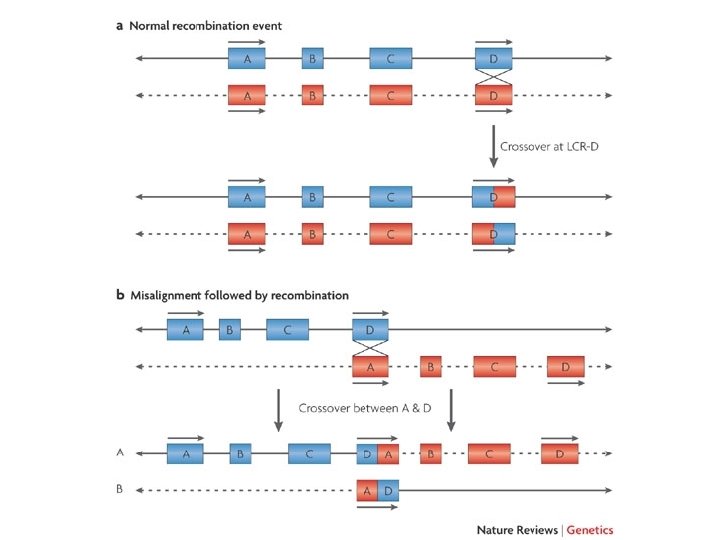
Nomenclatura delle delezioni • Le delezioni sono designate con la sigla del che segue i numeri dei nucleotidi a monte e a valle della delezione separatida un segno _ – 82_83 del (o 82_83 del. TG) indica una delezione di TG nella sequenza ACTTTGTGCC (dove A è il nucleotide 76) che diventa ACTTTGCC
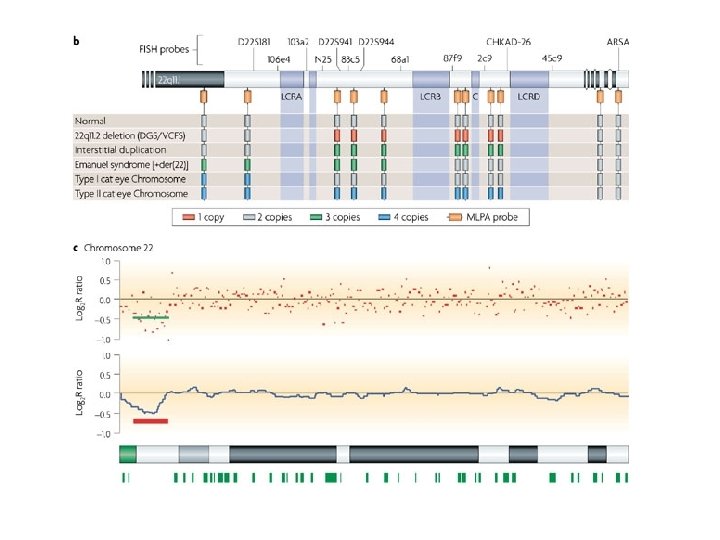
MLPA probes
Hybridization 1. 2. The MLPA probemix is added to denatured genomic DNA The two parts of each probe hybridise to adjacent target sequences
ligation 3. Probes are ligated by a thermostable ligase
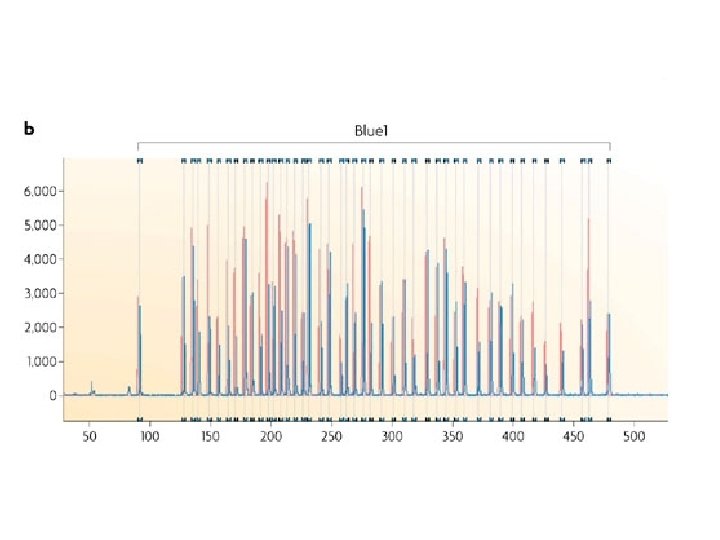
PCR amplification 4. A universal primer pair is used to amplify all ligated probes The PCR product of each probe has a unique length (130 480 bp)
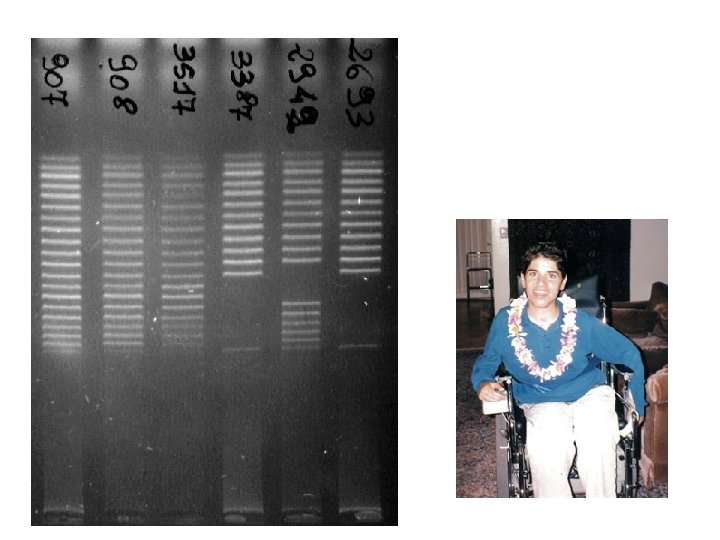
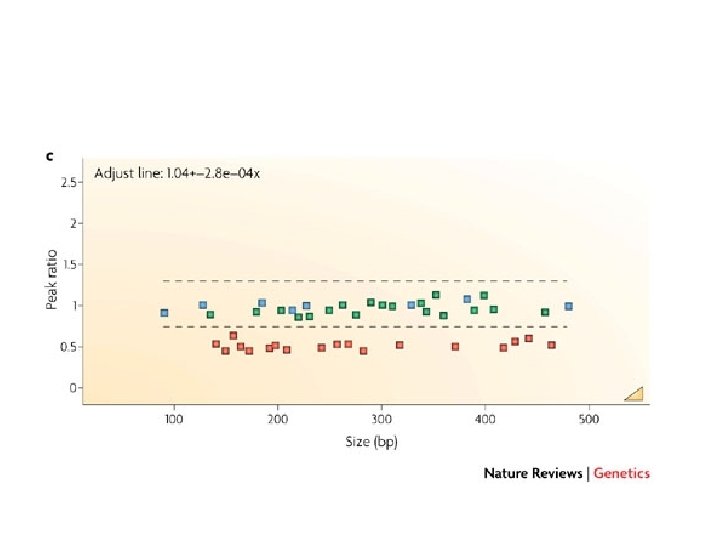
separation and quantification by capillary electrophoresis Each peak is the amplification product of a specific probe. Samples are compared to a control sample. A difference in relative peak height or peak area indicates a copy number change of the probe target sequence
detection of Chr X copy number X Male Female Triple X 283 bp 346 bp
MLPA discriminates sequences that differ in only a single nucleotide and can be used to detect known mutations Mismatch at the probe ligation site No ligation, no amplification product Perfect match Ligation of the two probe oligonucleotides Amplification product
MS-MLPA M M Methylated Target Denaturation and Multiplex probe hybridization Ligation and Digestion with methylation sensitive endonucleases Unmethylated Target M M Only undigested (methylated) and ligated probes are exponentially amplified