Nukleov kyseliny Struktura DNA a RNA Milada Tepl

 v buněčném jádře ribonukleová (RNA) v cytoplasmě a v")






 Vybírá správné aminokyseliny a umísťuje je do správného místa")


 Histonový komplex (8 nm) nukleosom")

 30")





 Syntéza DNA 3' Templátový Řetězec (DNA) T 5' T")








 3' Vedoucí řetězec Spustit animaci 1. Primasa 2. DNA-polymerasa")

 3' Vedoucí řetězec Spustit animaci 3. DNA-polymerasa I (exonukleasová")


n + d. NTP ⇄ (DNA)n + 1")
 Milada Teplá, Helena Klímová KUDCH,")



 U Nově syntetizovaný Řetězec (RNA)")




 neboli heterogenní")
 • nekódující sekvence")
 Milada Teplá, Helena Klímová KUDCH,")

 kombinací trojic nukleotidů: • některé aminokyselině přísluší")



 je navázána aminokyselina Antikodon jsou tři")


 t.")













. •")
. •")


- Slides: 76
Nukleové kyseliny Struktura DNA a RNA Milada Teplá, Helena Klímová KUDCH, PřF UK v Praze 2013
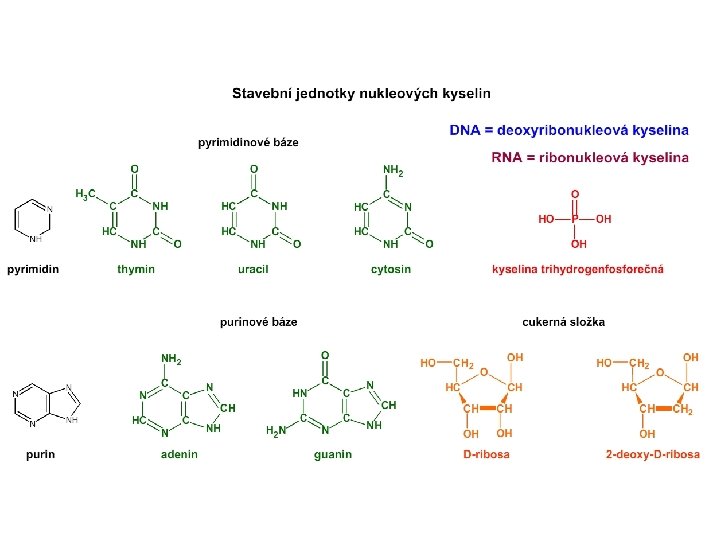
Typy nukleových kyselin deoxyribonukleová (DNA) v buněčném jádře ribonukleová (RNA) v cytoplasmě a v jadérku
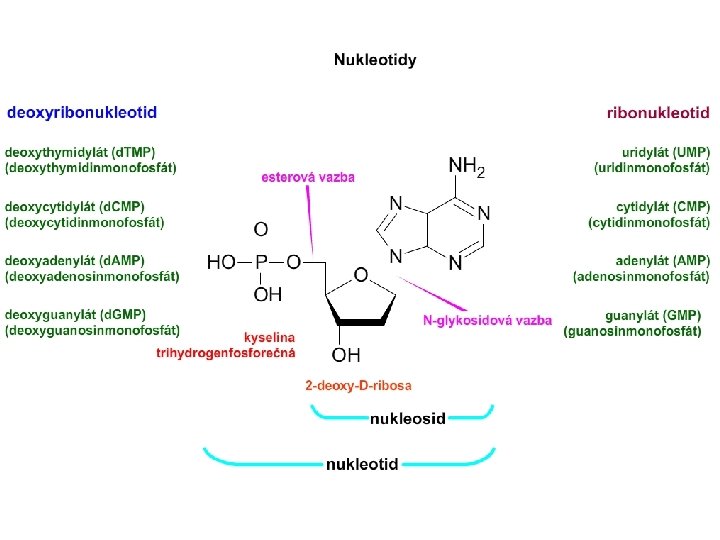
Mezi fosfátovou skupinou na 5. C a -OH skupinou na 3. C na pentose vzniká tzv. fosfodiesterová vazba Na povrchu je NA velmi silně záporně nabitá.
Dusíkaté báze se mohou mezi sebou pomocí vodíkových vazeb párovat
Toto komplementární párování bází umožňuje zaujmout energeticky nejvýhodnější konformaci
3' 5' DNA - pravotočivá šroubovice DNA je stočena do pravotočivé šroubovice, která je stočena ze dvou komplementárních vláken DNA. Řetězce v DNA jsou vůči sobě antiparalelní. 5' 3'
Typy DNA B DNA Pravotočivá. Zhruba 10 bází na závit. Báze tvořící pár leží vždy v jedné rovině. Dvoušroubovice DNA má na svém povrchu dva typy žlábků (malý a velký). A DNA Pravotočivá. 11 párů bází na závit. Z DNA Levotočivá. 12 párů bází na závit. Vytváří se, pokud se ve šroubovici objeví pravidelné opakování bází adeninu a thyminu. Prokaryotní DNA i DNA semiautonomních organel (např. mitochondrií) jsou kruhové.
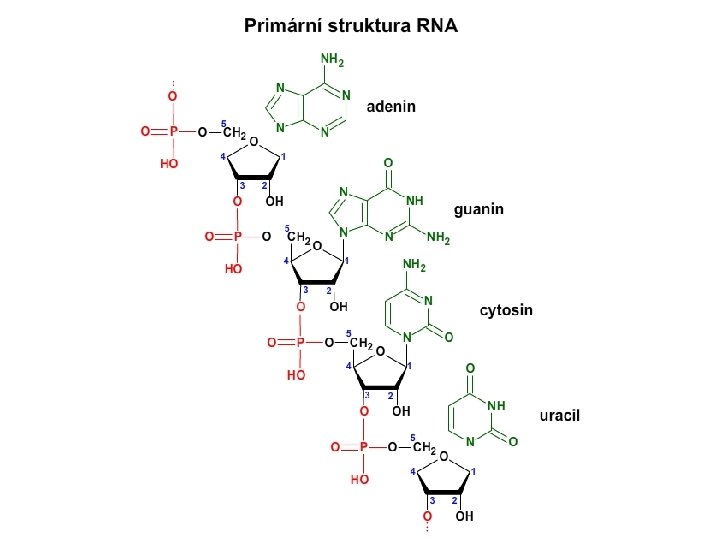
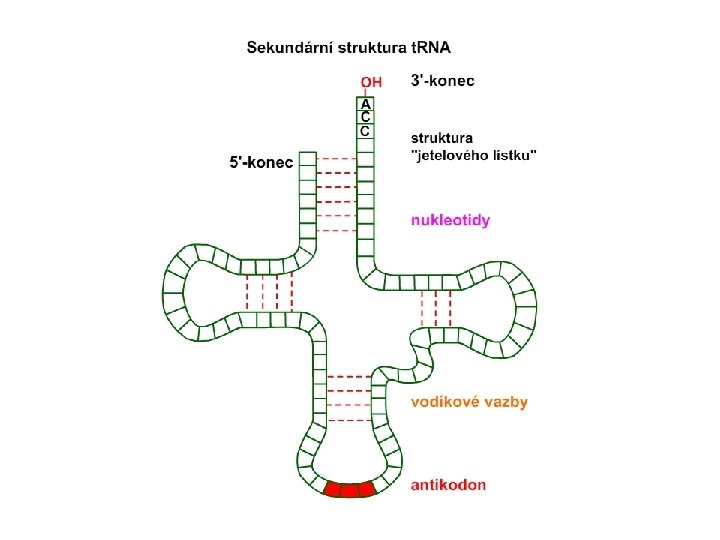
Molekula RNA Jednořetězcová. Primární struktura RNA Sekundární struktura t. RNA
Typy RNA t. RNA (transferová) Vybírá správné aminokyseliny a umísťuje je do správného místa na ribosomu, aby mohly být začleněny do rostoucího aminokyselinového řetězce. r. RNA m. RNA (ribosomální) (informační, mediatorová) Tvoří jádro ribosomů, na kterých je m. RNA překládána do proteinu. Vzniká přepisem genů kódujících aminokyselinovou sekvenci proteinů a její základní funkcí je řídit vznik proteinu. r. RNA a t. RNA vznikají přepisem genů nekódujících aminokyselinovou sekvenci proteinu. Jedná se o tzv. neinformační RNA.
Chromosom Milada Teplá, Helena Klímová KUDCH, PřF UK v Praze 2013
Chromosom Jádro lidské buňky – průměr cca 5 -8 μm DNA – cca 2 m V eukaryotních buňkách – molekuly DNA jsou asociovány se specifickými proteiny a sbaleny do chromosomů. Mitotické chromosomy • v době buněčného dělení (v mitose) • v kondenzovaném stavu (sbaleném) Interfázové chromosomy • značně rozvolněné a vypadají jako jedna dlouhá, tenká, vzájemně propletená vlákna
Interfázový chromosom – Korálková forma chromatinu (10 -nm vlákno) Histonový komplex (8 nm) nukleosom Spojníková DNA 10 nm Chromatin – komplex DNA a proteinů Histony – malé proteiny s vysokým obsahem kladně nabitých AMK (lys a arg)
Mitotický chromosom Před vstupem do mitosy dochází k sbalování chromatinu. • Chromatin se stáčí za vzniku tzv. solenoidu. Solenoid: 6 nukleosomů na závit 30 nm • 30 -nm vlákno je organizováno do smyček připojených k centrální ose, tzv. lešení (scaffold). • Solenoid se stáčí na chromatidu.
2 nm 10 nm Úsek dvoušroubovice DNA Korálková forma chromatinu (10 -nm vlákno) 30 nm 30 -nm vlákno 300 nm Rozvinutý úsek chromosomu 700 nm Kondenzovaný úsek chromosomu 1400 nm Úplný mitotický chromosom
Replikace DNA Milada Teplá, Helena Klímová KUDCH, PřF UK v Praze 2013
G G T C A T TA G C G G A T C C T G C T A A T C TA G G G C C CG A T T Nově syntetizovaný řetězec DNA G CG A C G A T A CG A C Templát (matrice) A A T C C T
Replikační počátky a replikační vidličky Celý proces replikace začíná v replikačních počátcích. Replikační počátek dvoušroubovice DNA Replikační počátky se zvětšují za vzniku tzv. replikačních vidliček. Replikační vidlička
Replikační vidličky: Helikasa = enzym, rozvíjí dvoušroubovicovou strukturu
Replikační vidličky: obousměrnost replikace V jednom replikačním počátku se vytvoří dvě replikační vidličky. Replikace je obousměrná. Replikační vidlička Směr replikace Začátek replikace
5' Nově syntetizovaný Řetězec (DNA) Syntéza DNA 3' Templátový Řetězec (DNA) T 5' T A A 3' T A A T 3'-konec 5'-konec G G C C Fosfodiesterová vazba 5' 3' 5'
Syntéza DNA: DNA-polymerasa • enzym • připojuje nukleotidy na 3'-konec rostoucího řetězce DNA • vznik fosfodiesterové vazby mezi 3'-OH skupinou řetězce a 5'-fosfátovou skupinou přidávaného nukleotidu. Nukleotidy vstupují do reakce jako energeticky bohaté deoxynukleosidtrifosfáty (např. d. ATP) – dodání energie polymerizační reakci. Většinu nové DNA polymerizuje DNA-polymerasa III. DNA je syntetizována ve směru 5' → 3'.
DNA-polymerasa 3' Spustit animaci 5' Primer Vedoucí řetězec 5' 3' 3' Deoxynukleosidtrifosfáty (d. ATP, d. GTP, d. TTP a d. CTP) Enzym: DNA-polymerasa III 5'
Syntéza DNA: Primasa DNA-polymerasa neumí začít syntetizovat nové vlákno. Primasa – enzym, dokáže spojit dva volné nukleotidy. Primasa syntetizuje krátké úseky RNA (cca 10 nukleotidů). Tyto úseky poskytují 3'-konec pro DNA-polymerasu. Slouží tedy jako primer pro syntézu DNA.
Primasa Enzym: Primasa Primer 5' 5' 3' 3' Enzym: DNA-polymerasa 5'
Asymetričnost replikační vidličky První řetězec je syntetizován podle templátu ve směru 3'→ 5' (Vzniká 5'→ 3' řetězec) Druhý řetězec je syntetizován podle templátu ve směru 5'→ 3' 3' (Vzniká 3'→ 5' řetězec). 5' 5' 3' 3' 5' Směr replikace 3' 5'
Asymetričnost replikační vidličky: Okazakiho fragmenty DNA-polymerasa syntetizuje nové vlákno pouze prodlužováním 3'-konce DNA (nový řetězec může vznikat jen ve směru 5'→ 3', neboli dle templátu 3'→ 5'). Je-li DNA syntetizována dle templátu 5'→ 3': - replikace probíhá diskontinuálně a „po zpátku“ - syntetizují se tzv. Okazakiho fragmenty 3' 5' 3' Směr replikace 5'
Asymetričnost replikační vidličky: vedoucí a váznoucí řetězec vedoucí řetězec 3' 5' 5' 3' 3' 5' „Po zpátku“ 3' opožďující se (váznoucí řetězec)
Asymetričnost replikační vidličky 3' 5' Vedoucí řetězec Váznoucí řetězec 3' 5' 5' 3' Okazakiho fragmenty 3' 5' 5' 5' 3' Směr pohybu replikační vidličky 5' 3' 3' 5' nejnověji nasyntetizovaná DNA
Váznoucí řetězec (I. část animace) 3' Vedoucí řetězec Spustit animaci 1. Primasa 2. DNA-polymerasa III 5' 5' 3' Okazakiho fragmenty 3' 5' Váznoucí řetězec
Váznoucí řetězec Opožďující řetězec je tvořen mnoha oddělenými úseky tzv. Okazakiho fragmenty (cca 1000 -2000 nukleotidů). Na vytvoření souvislého vlákna DNA z Okazakiho fragmentů jsou třeba tři enzymy: 1. DNA-polymerasa I (exonukleasová funkce) – odstraňuje RNA primery; 2. DNA-polymerasa I – nahrazuje RNA-primery DNA; 3. DNA-ligasa – spojí všechny úseky dohromady.
Váznoucí řetězec (II. část animace) 3' Vedoucí řetězec Spustit animaci 3. DNA-polymerasa I (exonukleasová funkce) 5' 5' 3' 4. DNA-polymerasa I 5. DNA-ligasa 3' 5' Váznoucí řetězec
Replikační aparát 5' 3' Vedoucí řetězec Svírací protein DNA-polymerasa Nově syntetizovaný řetězec Primasa Nový Okazakiho fragment 5' 3' Rodičovská DNA 3' 5' DNA-helikasa RNA-primer Vazebný protein pro udržení jednořetězcové Váznoucí řetězec struktury Okazakiho fragment
Replikace je proces semikonzervativní 5' 3' 3' 5' 5' 3'
Korektorská schopnost DNA-polymerasy DNA-polymerasa katalyzuje reakci: (DNA)n + d. NTP ⇄ (DNA)n + 1 + difosfát DNA-polymerasa je schopna hydrolyzovat DNA od 3'-konce (tzv. 3'→ 5' exonukleasová aktivita). Připojí-li se chybný nukleotid, vznikne nestabilní produkt, čímž dojde k posunutí rovnováhy ve směru výchozích látek. DNA-polymerasa je velice přesně párující enzym: udělá průměrně 1 chybu na 107 zreplikovaných párů bází.
Transkripce (první krok genové exprese: Od DNA k RNA) Milada Teplá, Helena Klímová KUDCH, PřF UK v Praze 2013
Transkripce a translace Jestliže buňka potřebuje nějaký konkrétní protein, je nukleotidová sekvence je v patřičné oblasti v chromosomu nejprve zkopírována do m. RNA. Tato RNA je využívána jako templát (předloha, matrice) pro tvorbu proteinů. transkripce DNA translace m. RNA Protein
Transkripce - úvod transkripce DNA m. RNA Transkripce: • gen se přepisuje do RNA • začíná rozvolňováním krátkého úseku DNA • • jeden z řetězců DNA slouží jako templát pro syntézu RNA Dvoušroubovice DNA
Komplementarita bází
3' 5' Syntéza RNA 5' 3' Templátový Řetězec (DNA) U Nově syntetizovaný Řetězec (RNA) Templátový Řetězec (DNA) A A U T Nově syntetizovaný Řetězec (RNA) 3'-konec A A T 5'-konec G G C 5' Fosfodiesterová vazba 3' C 5'
Vznik transkriptu Transkript - řetězec RNA vznikající transkripcí. Dochází k obnovení dvoušroubovicové struktury DNA a vytěsnění vlákna RNA – molekuly RNA jsou jednovláknové. Ribonukleosidtrifosfáty Transkript (m. RNA) Templát Dvoušroubovice DNA Směr transkripce
RNA-polymerasa katalyzuje připojování nukleotidů na 3'-konec rostoucího řetězce RNA za vzniku fosfodiesterové vazby mezi 3'-OH skupinou řetězce a 5'-fosfátovou skupinou přidávaného nukleotidu. RNA je syntetizována ve směru 5' → 3'. Pro syntézu RNA je využívána energie vznikající hydrolýzou ribonukleosidtrifosfátu (ATP, UTP, GTP a CTP). Vznik fosfodiesterové vazby 5' 3'
RNA-polymerasa Ribonukleosidtrifosfáty Templát pro syntézu RNA Rozvíjecí místo
Transkripce 3' Spustit animaci Transkript 5' Ribonukleosidtrifosfát 5' 3' Templát 3' 5'
Posttranskripční úpravy RNA u eukaryot Transkripcí vzniká nejprve primární transkript (Pre-m. RNA) neboli heterogenní jaderná RNA (hn. RNA) Ta se před opuštěním z jádra dále upravuje (podléhá posttranskripčním úpravám). Upravená m. RNA je transportována do cytoplasmy a na ribosomech je překládána na proteiny (translace). Posttranskripční úpravy transkripce DNA (v jádře) Pre-m. RNA (v jádře) translace m. RNA (vznik v jádře, transport do cytoplasmy) Protein
Exony a introny Eukaryotní DNA obsahuje: • kódující sekvence (tzv. exony) • nekódující sekvence (tzv. introny) Sestřih (RNA splicing): • Introny z Pre-m. RNA jsou odstraňovány „sestřihovými“ enzymy • exony jsou spojeny dohromady m. RNA Exony Introny 2. Odštěpení intronů a spojení exonů 1. Přiblížení obou konců intronů
Translace (druhý krok genové exprese: Od RNA k proteinu) Milada Teplá, Helena Klímová KUDCH, PřF UK v Praze 2013
Genetický kód a kodon Z funkční m. RNA je informace použita pro syntézu proteinu. Genetický kód: pravidla, kterými se řídí přenos z DNA do aminokyselinové sekvence (prostřednictvím m. RNA). V genetickém kódu platí konvence, že 5'-konec m. RNA je zapisován vlevo! Sekvence nukleotidů m. RNA je čtena po trojicích – po kodonech. G G C C A G U U A U G C G U m. RNA A C A A 5' kodon 3'
Genetický kód Dohromady lze vytvořit 64 (43) kombinací trojic nukleotidů: • některé aminokyselině přísluší i několik tripletů (např. CCU, CCA, CCG, CCC je prolin) • jednomu tripletu přísluší nanejvýš jedna aminokyselina. Genetický kód je téměř univerzální pro všechny organismy. G G C C A G U U A U G C G U m. RNA A C A A 5' kodon 3'
Genetický kód V principu může být m. RNA překládána ve všech třech čtecích rámcích podle toho, u kterého nukleotidu translace začne. Avšak jen v jednom čtecím rámci vzniká požadovaný protein. Val Thr 5' 5' 5' Ala Lys Val Stop kodon G G C C A G U U A U G C G U m. RNA A C A A Gln Leu Arg Pro Ser G G C C A G U U A U G G U C m. RNA A C A A Ser Gly Cys 3' Gln Leu G G C C A G U U A U G C G U m. RNA A C A A 3' 3'
Genetický kód – všech 64 možných kombinací U C První nukleotid A Druhý nukleotid U A Terminační kodony G UUU fenyalanin UCU serin UAU tyrosin UGU cystein U UUC fenyalanin UCC serin UAC tyrosin UGC cystein C UUA leucin UCA serin UAA stop kodon A UUG leucin UCG serin UAG stop kodon UGA stop kodon (selenocystein) UGG tryptofan G CUU leucin CCU prolin CAU histidin CGU arginin U CUC leucin CCC prolin CAC histidin CGC arginin C CUA leucin CCA prolin CAA glutamin CGA arginin A CUG leucin CCG prolin CAG glutamin CGG arginin G AUU isoleucin ACU threonin AAU asparagin AGU serin U AUC isoleucin ACC threonin AAC asparagin AGC serin C AGA arginin A AUA isoleucin G C Iniciační kodon ACA threonin AAA lysin AUG methionin ACG threonin AAG lysin AGG arginin G GUU valin GCU alanin GAU kyselina asparagová GGU glycin U GUC valin GCC alanin GAC kyselina asparagová GGC glycin C GUA valin GCA alanin GAA kyselina glutamová GGA glycin A GUG valin GCG alanin GAG kyselina glutamová GGG glycin G Třetí nukleotid
t. RNA Kodony v m. RNA nerozpoznávají přímo aminokyseliny, které specifikují. Translace m. RNA do proteinu závisí na t. RNA, která se spáruje s kodonem v m. RNA. Na t. RNA je navázána aminokyselina (aminoacyl-t. RNA). Touto částí váže příslušné aminokyseliny Touto částí se páruje s kodonem v m. RNA
Aminoacyl-t. RNA Na 3'-konec (vždy končí sekvencí CCA) je navázána aminokyselina Antikodon jsou tři nukleotidy komplementární ke kodonu v m. RNA
Struktura aminoacyl-t. RNA přibližně 80 nukleotidů dlouhé Skutečný L-tvar t. RNA Struktura jetelového listu
Aminoacyl-t. RNA-synthetasa - enzym, rozpozná a připojí správnou aminokyselinu k t. RNA Kodon pro tryptofan je UGG – antikodon je ACC Aminokyselina (Tryptofan) t. RNA A C C Aminoacyl-t. RNA-synthetasa
Aminoacyl-t. RNA-synthetasa Reakce katalyzovaná aminoacyl-t. RNA-synthetasou vyžaduje dodání energie hydrolýzou ATP. Aminokyselina (Tryptofan) t. RNA ATP A C C Aminoacyl-t. RNA-synthetasa AMP + 2 Pi Vazba aminokyseliny k t. RNA A C C
Aminoacyl-t. RNA-synthetasa Při této reakci vzniká vysokoenergetická vazba mezi t. RNA a AMK. Energie je využita pro tvorbu kovalentní vazby mezi rostoucím polypeptidovým řetězcem a nově navázanou aminokyselinou. Aminokyselina (Tryptofan) Makroergická vazba t. RNA ATP A C C Aminoacyl-t. RNA-synthetasa AMP + 2 Pi Vazba aminokyseliny k t. RNA A C C Vazba kodonu k antikodonu 5' A C C Párování U G G m. RNA bází 3'
Translace probíhá na ribosomech E-místo P-místo A-místo Velká ribosomální jednotka Vazebné místo pro m. RNA E P A Malá ribosomální jednotka Malá podjednotka zodpovídá za nasednutí t. RNA na kodon m. RNA. Velká podjednotka katalyzuje vznik peptidové vazby mezi aminokyselinou a polypeptidovým řetězcem. Ribosom se pohybuje podél m. RNA, překládá sekvenci m. RNA do aminokyselinové sekvence za použití t. RNA.
Met Iniciace translace Začátek na iniciačním kodonu AUG Pro iniciaci je třeba iniciační t. RNA, s navázaným methioninem (u bakterií iniciační formyl-methionin). t. RNA s Met Malá ribosomální podjednotka 5' U eukaryot: iniciační t. RNA je připojená k malé ribosomální jednotce. AUG 3' m. RNA
Iniciace translace malá podjednotka se váže na 5'-konec m. RNA pohybuje se ve směru 5' → 3' a hledá kodon AUG, který je rozpoznán antikodonem iniciační t. RNA. Met 5' AUG 3' m. RNA
Iniciace translace E P A malá podjednotka se váže na 5'-konec m. RNA pohybuje se ve směru 5' → 3' a hledá kodon AUG, který je rozpoznán antikodonem iniciační t. RNA. Po rozpoznání iniciačního kodonu se připojí velká ribosomální podjednotka. Velká ribosomální podjednotka Met E 5' P A Iniciační t. RNA se váže do P-místa, prodlužování řetězce může ihned začít navázáním druhé t. RNA s aminokyselinou do A-místa. AUG 3' m. RNA
Iniciace translace aa 2 malá podjednotka se váže na 5'-konec m. RNA pohybuje se ve směru 5' → 3' a hledá kodon AUG, který je rozpoznán antikodonem iniciační t. RNA. Po rozpoznání iniciačního kodonu se připojí velká ribosomální podjednotka. Met E 5' P A Iniciační t. RNA se váže do P-místa, prodlužování řetězce může ihned začít navázáním druhé t. RNA s aminokyselinou do A-místa. AUG 3' m. RNA
Iniciace translace V dalším kroku dochází ke vzniku peptidové vazby mezi methioninem a přicházející aminokyselinou (aa 2). Met E 5' P AUG aa 2 A m. RNA 3'
Iniciace translace V dalším kroku dochází ke vzniku peptidové vazby mezi methioninem a přicházející aminokyselinou (aa 2). Ribosom se posune o 3 nukleotidy podél m. RNA. t. RNA bez navázané aminokyseliny se uvolní z E-místa a t. RNA z A-místa se přenese do P-místa. Met aa 2 E 5' P AUG A m. RNA 3'
Elongace translace Neustále opakován tříkrokový cyklus: • První krok: aminoacyl-t. RNA je navázána do A-místa. aa 4 NH 2 aa 1 aa 2 E 5' aa 3 P A m. RNA 3'
Elongace translace Neustále opakován tříkrokový cyklus: • První krok: aminoacyl-t. RNA je navázána do A-místa. • Druhý krok: vznik peptidové vazby. NH 2 aa 1 aa 2 E 5' aa 3 P aa 4 A m. RNA 3'
Elongace translace Neustále opakován tříkrokový cyklus: • První krok - aminoacyl-t. RNA je navázána do A-místa. • Druhý krok - vznik peptidové vazby. • Třetí krok - ribosom se posune o 3 nukleotidy. - t. RNA bez navázané AMK se uvolní z Emísta. - t. RNA z A-místa se přenese do P-místa. NH 2 aa 1 aa 2 aa 3 aa 4 E 5' P A m. RNA 3'
Elongace translace - aminoacyl-t. RNA je navázána do A-místa. • První krok • Druhý krok - vznik peptidové vazby. • Třetí krok - ribosom se posune o 3 nukleotidy. - t. RNA bez navázané AMK se uvolní z Emísta. - t. RNA z A-místa se přenese do P-místa. aa 5 NH 2 aa 1 aa 2 aa 3 E 5' aa 4 aa 5 P A m. RNA 3'
Rostoucí peptidový řetězec Elongace translace NH 2 aa 1 aa 2 aa 3 EPA 5' m. RNA je překládána ve směru 5' → 3' nejprve vzniká N-konec proteinu (polypeptidový řetězec roste směrem od N-konce k C-konci). aa 4 NH 2 m. RNA 3' aa 1 aa 2 aa 3 aa 4 EPA 5' NH 2 m. RNA aa 1 aa 2 aa 3 aa 4 3' Posun o tři nukleotidy EPA Celý cyklus všech tří kroků je opakován dokud ribosom nenarazí na stop-kodon. 5' m. RNA 3' aa 5 NH 2 aa 1 aa 2 aa 3 aa 4 EPA 5' NH 2 t. RNA m. RNA 3' aa 1 aa 2 aa 3 aa 4 aa 5 EPA 5' m. RNA 3'
Terminace translace Konec proteinu je signalizován přítomností stop kodonu (UAA, UAG nebo UGA). • do A-místa se váže uvolňovací faktor • místo AMK se váže molekula vody • uvolní se C-konec polypeptidového řetězce z t. RNA v P-místě Uvolňovací faktor NH 2 aa 1 aa 2 aa 3 aa 4 E 5' aa 5 P A UAA m. RNA 3'
Terminace translace Konec proteinu je signalizován přítomností stop kodonu (UAA, UAG nebo UGA). • do A-místa se váže uvolňovací faktor • místo AMK se váže molekula vody • uvolní se C-konec polypeptidového řetězce z t. RNA v P-místě H 2 O NH 2 aa 1 5' aa 2 aa 3 aa 4 aa 5 E P Protein se uvolňuje do cytoplasmy. COOH A UAA m. RNA 3'
Terminace translace • m. RNA se odpojí od ribosomu • dojde k disociaci obou podjednotek ribosomu 5' E P A UAA m. RNA 3'
Použitá literatura ALBERTS, B. a kol. Základy buněčné biologie. Ústí nad Labem: Espero Publishing, 1997. NEČAS, O. a kol. Obecná biologie pro lékařské fakulty. Jinočany: Nakladateství H&H, 2000. KUBIŠTA, V. Buněčné základy životních dějů. Praha: Scientia, 1998. Obrázky: Milada Teplá (v MS powerpoint, Adobe flash)