Recap eukaryotes have 3 nuclear RNA polymerases which
































 of RNA polymerase II CTD facts: - unique to RNA")














 Clamp Tail F. Asturias Srb")













- Slides: 63
Recap • eukaryotes have 3 nuclear RNA polymerases, which transcribe unique sets of genes • RNA pol II transcribes protein coding genes and must respond to and integrate a diverse set of signals in order to regulate expression of >25 k genes • in vitro transcription systems for pol II show accurate initiation • gene specific regulators in euks have separable DNA binding and activation domains, the role of the DNA binding domain is to tether the activation domain near the promoter • activation domains have no clear distinguishing structural or sequence features that indicate their mechanism of action • squelching experiments indicate that activators compete for some limiting factor (not the polymerase) • TFIID and holoenzyme hypotheses may explain activator function
activator interference or squelching activator B activator A UAS hypothesis? TATA box
what is the limiting target of activators? 1. Eukaryotic activators do not bind to RNA pol II polymerase and therefore do not directly recruit polymerase to promoters. 2. Activators may, however, indirectly recruit RNA polymerase by recruiting factors (often called co-activators) that serve as a physical bridge between activator and polymerase. ‘TFIID hypothesis’ ‘Holoenzyme hypothesis’
Isolation of coactivators associated with the TATA-binding protein that mediate transcriptional activation Dynlacht, Hoey, Tjian, Cell 1991 Robert Tjian in vitro transcription reactions assembled from partially purified basal transcription factors -pol II ~90% pure -general factors <1% pure when assaying basal transcription (no activator present) in vitro, recombinant TBP can substitute for TFIID
Isolation of coactivators associated with the TATA-binding protein that mediate transcriptional activation Dynlacht, Hoey, Tjian, Cell 1991 recombinant TBP cannot substitute for TFIID when assaying activated transcription in vitro
Isolation of coactivators associated with the TATA-binding protein that mediate transcriptional activation Dynlacht, Hoey, Tjian, Cell 1991 TBP, the TATA-binding protein is small, but glycerol gradient sedimentation and gel filtration chromatography indicates that TFIID is very large
Isolation of coactivators associated with the TATA-binding protein that mediate transcriptional activation Dynlacht, Hoey, Tjian, Cell 1991 the coactivator activity can be separated from TBP by ion exchange chromatography of TFIID in the presence of urea TBP and TBP-associated factors are required for activated transcription
RNA Polymerase II Transcription Machinery Number of subunits Pol II 12 GTFs TFIID TFIIB TFIIE TFIIH TFIIF TFIIA* Mediator TBP TAFs* 1 12 1 2 9 2 3 22
Assembly of recombinant TFIID reveals differential coactivator requirements for distinct transcriptional activators Chen et al. , Cell 1994 had cloned and expressed most of the TAFs worked out methods for reconstitution of complex entirely from recombinant proteins
Assembly of recombinant TFIID reveals differential coactivator requirements for distinct transcriptional activators Chen et al. , Cell 1994
Assembly of recombinant TFIID reveals differential coactivator requirements for distinct transcriptional activators Chen et al. , Cell 1994 TAFII 150 and TAFII 60 are sufficient for activation by NTF-1
Assembly of recombinant TFIID reveals differential coactivator requirements for distinct transcriptional activators Chen et al. , Cell 1994 NTF-1 activation domain peptide on beads TAFII 150 and TAFII 60 are specifically retained
Assembly of recombinant TFIID reveals differential coactivator requirements for distinct transcriptional activators Chen et al. , Cell 1994
The ‘TFIID hypothesis’ 1. TAFs provide surfaces for the interaction of TFIID with activators. 2. TFIID recruits polymerase in vitro assays suggest specific activator-TAF contacts predictions?
Yeast TAFII 145 functions as a core promoter selectivity factor, not a general coactivator Walker et al. , Cell, 1997 Shen and Green, Cell, 1997 poly. A+ RNA levels are largely unaffected by inactivation of TAFs cell cycle regulated genes appear to be TAF-dependent
Yeast TAFII 145 functions as a core promoter selectivity factor, not a general coactivator Walker et al. , Cell, 1997 Shen and Green, Cell, 1997 TAFII 145 dependence tracks with the core promoter, not the UAS!
Transcriptional activation via enhanced preinitiation complex assembly in a human cell-free system lacking TAFIIs Oelgeschlager et al. , 1998 western blot demonstrating depletion of TAFIIs in vitro transcription shows that - transcription is abolished in the TFIID depleted extract - TBP is sufficient to restore activated transcription - 4 different activators were tested no transcription after depletion of TFIID and TAFs
Conclusions: 1. Several activators can activate transcription in vitro in the absence of TAFs. 2. Not all transcription depends on TAFs in vivo. (based on analysis of yeast TAF mutants) 3. Some TAFs may assist in recognition of the core promoter (rather than transmitting regulatory information associated with upstream factors). 4. TAFs and alternative TBPs may specify selection of particular core promoters.
What is the Limiting Target of Activators? activator B activator A UAS TATA box
A novel mediator between activator proteins and the RNA polymerase II transcription apparatus (Kelleher et al. 1990)
A novel mediator between activator proteins and the RNA polymerase II transcription apparatus (Kelleher et al. 1990) Gal 4 -VP 16 TATA X UASGAL 10 UAS(d. A-d. T)2 autoinhibition TATA activator interference UASGAL 10 UAS(d. A-d. T)2
A novel mediator between activator proteins and the RNA polymerase II transcription apparatus (Kelleher et al. 1990) Gal 4 -VP 16 Yeast nuclear extract in 50 m. M (NH 4)2 SO 4 X UAS(d. A-d. T)2 TATA 400 m. M Elu DEAE column 50 m. M 400 m. M (NH 4)2 SO 4 50 m. M FT the 400 m. M fraction overcomes squelching by Gal 4 -VP 16
A novel mediator between activator proteins and the RNA polymerase II transcription apparatus (Kelleher et al. 1990) Potential explanations? -column fraction has activator for the template -something in column is binding/sequestering Gal 4 -VP 16 -general stimulatory effect -fraction contains some limiting basal factor
A novel mediator between activator proteins and the RNA polymerase II transcription apparatus (Kelleher et al. 1990) Potential explanations? -column fraction has activator for the template -no, it doesn’t squelch a Gal 4 template -something in column is binding/sequestering Gal 4 -VP 16 -no, activation by Gal 4 -VP 16 is not disrupted -general stimulatory effect -no, activation depends upon Gal 4 -VP 16 -fraction contains some limiting basal factor -no, adding them back does not overcome squelching
Squelching in vitro: interpretation autoinhibition UASG TATA hypothetical target of activators UASG TATA excess Gal 4 -VP 16 activator interference UASd. A-d. TTATA
A mediator required for activation of RNA polymerase II transcription in vitro Flanagan et al. , Nature, 1991 response to activators is lost during purification of general factors, but basal transcription (0 ug Gal 4 -VP 16) is unchanged mediator fraction restores activator response in a purified in vitro transcription system
A mediator required for activation of RNA polymerase II transcription in vitro Flanagan et al. , Nature, 1991 general transcription factors do not have mediator activity
A mediator required for activation of RNA polymerase II transcription in vitro Flanagan et al. , Nature, 1991 Gcn 4 squelches Gal 4 -VP 16 squelches Gcn 4 squelching is observed with the mediator fraction
A multiprotein mediator of transcriptional activation and its interaction with the C-terminal repeat domain of RNA polymerase II (Kim et al. Cell, 1994) -Srb 5 IP
A multiprotein mediator of transcriptional activation and its interaction with the C-terminal repeat domain of RNA polymerase II (Kim et al. Cell, 1994) components of holo-RNA polymerase II: -12 polymerase subunits -3 TFIIF subunits -SRB proteins -Gal 11, Sug 1
RNA polymerase II Transcription Machinery Number of subunits Pol II 12 GTFs TFIID TFIIB TFIIE TFIIH TFIIF TFIIA* Mediator TBP TAFs* 1 12 1 2 9 2 3 22
Crystal Structure of Yeast RNA Polymerase II at 2. 8 Å Resolution (Cramer et al, 2001) CTD
the carboxy-terminal domain (CTD) of RNA polymerase II CTD facts: - unique to RNA pol II - the CTD is not required for transcription in vitro -the CTD is essential for life Heptapeptide of the CTD (52 repeats in mammalian Rpb 1, 27 in yeast) -the CTD is subject to a cycle of phosphorylation at serines 2 and 5 - may be simultaneously phosphorylated at Ser 2, 5
A multiprotein mediator of transcriptional activation and its interaction with the C-terminal repeat domain of RNA polymerase II (Kim et al. Cell, 1994) an anti-CTD antibody separates mediator from RNA polymerase II
An RNA polymerase II holoenzyme responsive to activators Koleske and Young, Nature, 1994 previously: • CTD truncation mutations limit the response to activators • isolated srb mutation as suppressors of CTD truncation mutations in yeast • purified a complex of Srb proteins with pol II and basal factors “RNA polymerase II holoenzyme” Srb proteins copurify with RNA pol II
An RNA polymerase II holoenzyme responsive to activators Koleske and Young, Nature, 1994 previously: • CTD truncation mutations limit the response to activators • isolated srb mutation as suppressors of CTD truncation mutations in yeast • purified a complex of Srb proteins with pol II and basal factors “RNA polymerase II holoenzyme” Srb proteins copurify with RNA pol II holoenzyme supports activator dependent transcription in vitro
Transcriptional activation via enhanced preinitiation complex assembly in a human cell-free system lacking TAFIIs Oelgeschlager et al. , 1998 depletion of Srb 7 decreased transcription response to activator
The Med proteins of yeast and their function through the RNA polymerase II carboxy-terminal domain Myers et al. , Genes & Dev. , 1998 purified mediator binds the CTD in vitro
The Med proteins of yeast and their function through the RNA polymerase II carboxy-terminal domain Myers et al. , Genes & Dev. , 1998 the CTD is required for mediator activated transcription in vitro
a cycle of CTD modification during transcription TFIIH phosphorylates Ser 5 at initiation P-TEFb phosphorylates Ser 2 during elongation Ser 5 -P Ser 2 -P CTD phosphorylation: Pre-initiation/escape elongation
a cycle of CTD modification during transcription stage in transcription cycle pre-initiation early elongation phospho Ser 2, 5 later elongation CTD repeat=YS 2 PTS 5 PS
the CTD phosphorylation cycle coordinates diverse events during transcription mediator stage in transcription cycle pre-initiation capping enzyme early elongation later elongation splicing and poly. A factors
Association of an activator with an RNA polymerase II holoenzyme Hengartner et al. , 1995, Genes & Dev. holoenzyme is retained on a GST-VP 16 column a mutation that abolishes activation by VP 16 also abolishes holoenzyme binding
Activation domain-mediator interactions promote transcription preinitiation complex assembly on promoter DNA Cantin et al. , PNAS 2003 adenovirus E 1 A protein activates transcription of early genes by pol II E 1 A binds the Sur 2 subunit of mediator in vitro and associates with mediator in vivo E 1 A mutations that prevent activation also disrupt Sur 2 binding sur 2 -/- ES cells: -all other mediator subunits still in the complex -E 1 A doesn’t activate -several other activators still work
Activation domain-mediator interactions promote transcription preinitiation complex assembly on promoter DNA Cantin et al. , PNAS 2003 activation by E 1 A and Elk 1 in vitro requires Sur 2 no effect on activation by VP 16
Activation domain-mediator interactions promote transcription preinitiation complex assembly on promoter DNA Cantin et al. , PNAS 2003 Sur 2 is required for binding of mediator and GTFs to E 1 A-bound promoters Nuc. extract 5 x G 4 -act wash
Holoenzyme Hypothesis Mediator serves as a physical bridge between RNA pol II and activators by which activators recruit polymerase to the promoter. Mediator Holoenyme TFIIE TFIIF activator Pol II TFIIH TBP TFIIB PIC
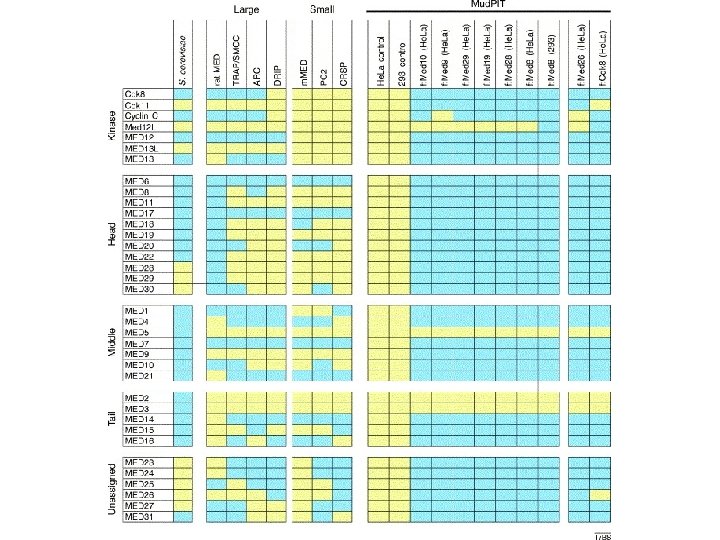
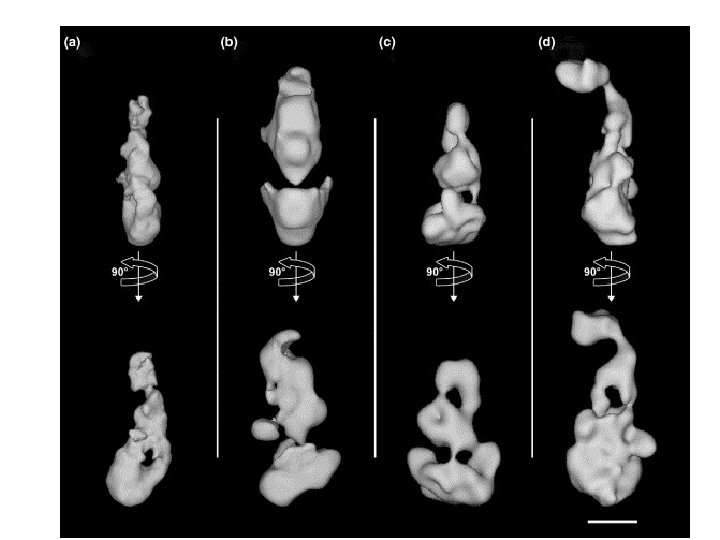
Mediator Bound to RNA Polymerase II (Single Particle analysis) Clamp Tail F. Asturias Srb 2 Srb 4 Head Srb 5 Head Srb 6 Med 6 Rgr 1 Med 8 Srb 7 Med 11 Med 1 Middle Med 4 Middle Med 7 Rox 3 Nut 1 Sin 4 Nut 2 Med 2 Tail Cse 2 Med 3 Gal 11
2 conformations of mediator
Holoenzyme Hypothesis Mediator serves as a physical bridge between RNA pol II and activators by which activators recruit polymerase to the promoter. Mediator Holoenyme TFIIE TFIIF activator Pol II TFIIH TBP TFIIB PIC
mediator summary 1. Of the 20 mediator subunits in yeast, 13 had been identified previously in genetic screens for factors affecting transcription. 1. 2. 11 mediator subunits are essential for life 2. 3. Mediator appears to required for all pol II transcription (a general factor? ) 4. Homologs for almost all Mediator subunits observed in fungi, plant and metazoan genomes. 1. 5. Strong structural similarity observed between mediator complexes of yeast, mice, humans 2. 6. In some cases, activators have been shown to contact specific mediator subunits and disruption of these contacts disrupts transcription
Holoenzyme Hypothesis Mediator serves as a physical bridge between RNA pol II and activators by which activators recruit polymerase to the promoter. Mediator Holoenyme TFIIE TFIIF activator Pol II TFIIH TBP TFIIB PIC predictions of model?
Gene activation by recruitment of the RNA polymerase II holoenzyme Farrel et al. , Genes and Dev. , 1996 recruitment of the mediator is sufficient for activated transcription
Association of the Mediator complex with enhancers of active genes Kuras et al. , PNAS 2003 Mediator Binding mediator binding did not depend on pol II binding or the TATA boxes TATA GAL 10 TATA enhancer GAL 1
The Swi 5 activator recruits the Mediator complex to the HO promoter without RNA polymerase II Bhoite et al. , Genes & Dev. , 2001 Mediator can be recruited to genes independently of Pol II, GTFs and transcription -often, recruitment is to enhancers rather than core promoter
Mediator as a general transcription factor Takagi and Kornberg, JBC, 2005 purified mediator from WT and srb 4 ts strains performed in vitro transcription reactions in the absence of activators “basal transcription”
Mediator as a general transcription factor Takagi and Kornberg, JBC, 2005 1. mediator behaves like a general transcription factor 2. temp. shift experiment show that it is required prior to initiation
Mediator as a general transcription factor Takagi and Kornberg, JBC, 2005 Srb 4 ts E(30°C) excess RNA pol II or basal factors cannot complement the transcription defect of the srb 4 ts mediator preparation suggests that mediator can act after recruitment of Pol II and general factors
Some TAFs function in promoter recognition TFIIB TBP TAFs TATA Inr DPE (Verrijzer et al, 1995)
New Model: TAFs function in core promoter recognition Are all TAFs devoted to promoter recognition? Goodrich et al, (1996)
TBP and TAF homologs may mediate tissue specific gene expression patterns in differentiated cells TRF=TBP related factor Reina JH, Hernandez N. Genes Dev. 2007