CARATTERISTICHE GENERALI DEI VIRUS ORGANIZZAZIONE NON CELLULARE MOLTIPLICAZIONE





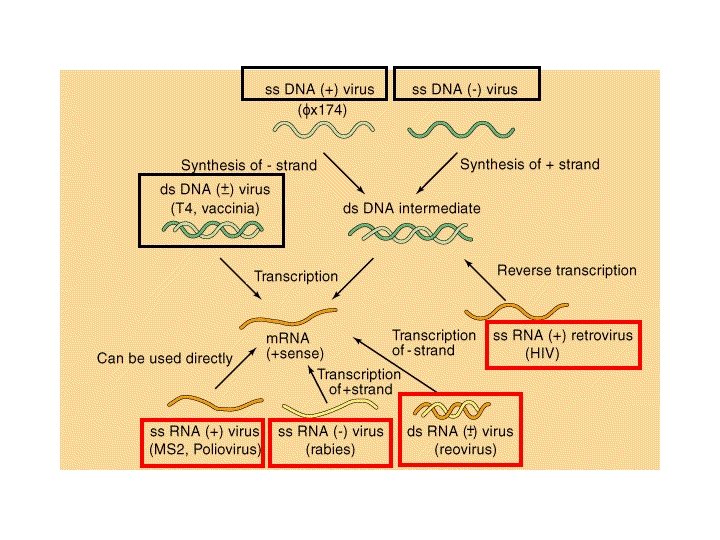
 HIV")


 litici")
 litici")









 temperati se alcuni geni del DNA virale vengono")






- Slides: 37
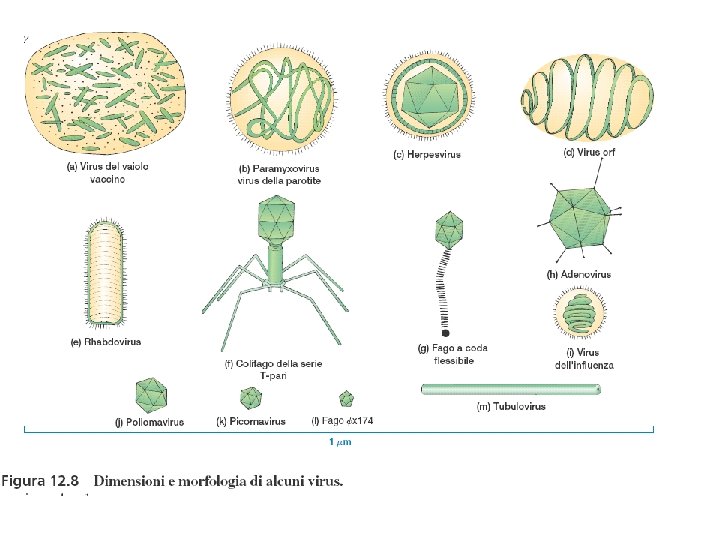
CARATTERISTICHE GENERALI DEI VIRUS • ORGANIZZAZIONE NON CELLULARE • MOLTIPLICAZIONE • PRESENZA DI UN UNICO TIPO DI ACIDO NUCLEICO • ASSENZA DI METABOLISMO PROPRIO MORFOLOGIA DEI VIRUS • ELICOIDALI • ICOSAEDRICI • RIVESTITI • COMPLESSI
VIRUS con morfologia elicoidale
VIRUS con morfologia icosaedrica
VIRUS rivestiti
VIRUS con morfologia complessa virus vaccino (vaiolo bovino) HIV
PFU = unità formanti placche
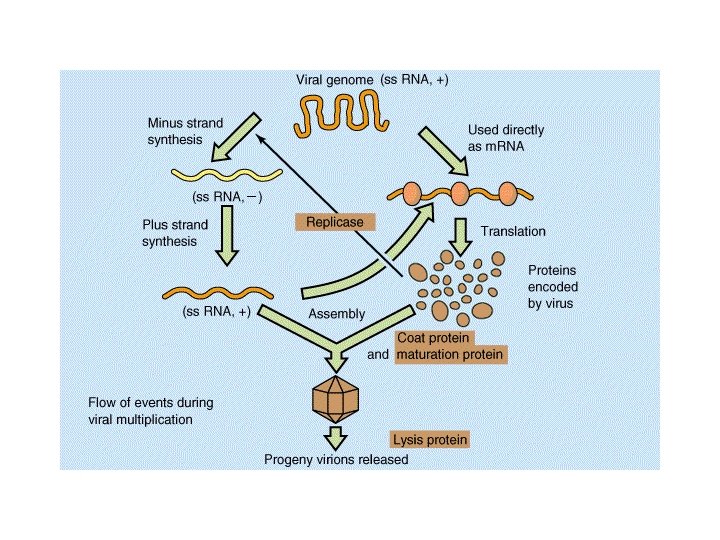
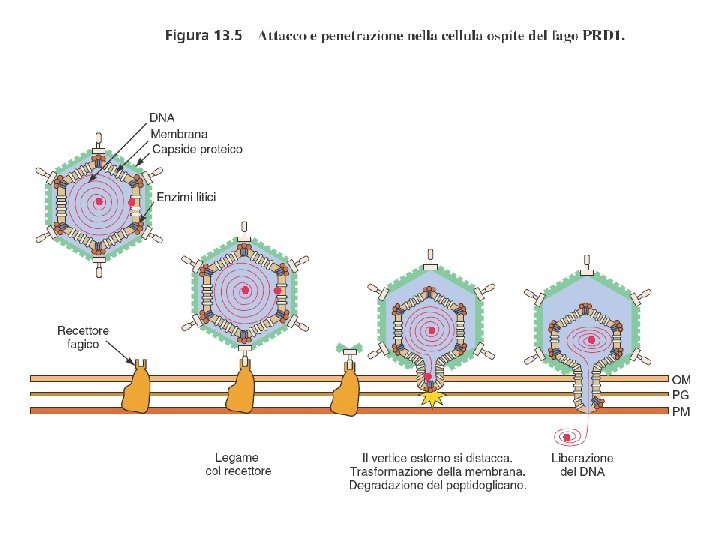
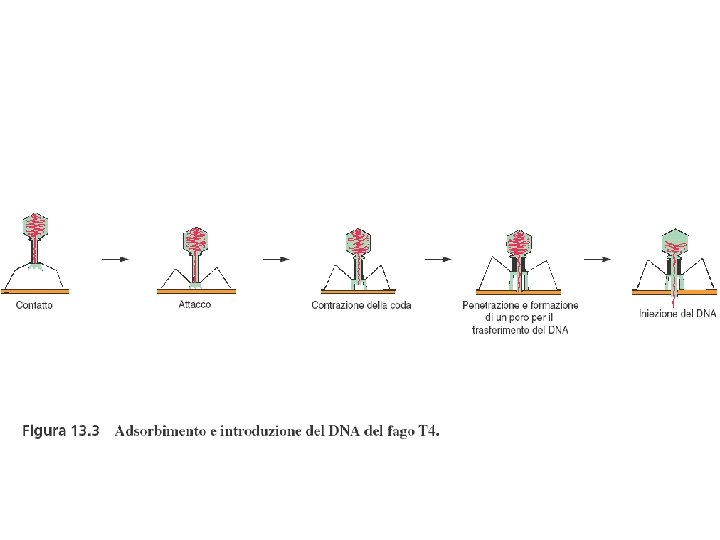
Meccanismo di infezione dei batteriofagi (fagi) litici
Meccanismo di infezione dei batteriofagi (fagi) litici
Le fasi della moltiplicazione dei batteriofagi 1. Adsorbimento 2. Iniezione dell’acido nucleico 3. Fase replicativa precoce 4. Replicazione del genoma virale 5. Sintesi delle proteine capsidiche 6. Assemblaggio del capside e impacchettamento del genoma virale 7. Rilascio dei virus maturi (lisi)
Curva di crescita a ciclo unico Numero dei batteriofagi periodo di latenza periodo di crescita plateau periodo di eclisse volume di scoppio = N. fagi al plateau N. fagi latenza Tempo (minuti) periodo di eclisse: periodo durante il quale non è possibile rilevare la presenza di particelle virali infettive neanche all’interno dei batteri periodo di latenza: periodo di sviluppo dei fagi nel batterio infettato. I fagi non sono ancora liberati all’esterno. Nella fase finale sono presenti particelle mature nel batterio. periodo di crescita: le cellule vanno incontro a lisi liberando i fagi infettanti. Quando tutti i batteri infettati saranno lisati si raggiunge il plateau
LATENZA: fase in cui i virioni sono ancora all’interno della cellula infettata ECLISSE: fase in cui i virioni all’interno della cellula non sono in grado di infettare. La latenza continua fino allo scoppio delle cellule infettate NUMERO DI VIRIONI MATURI NUMERO DI VIRUS LIBERI
La replicazione del DNA di T 4 concatenamero Dopo diversi cicli di replicazione si vengono a formare concatenameri di lunghezza considerevole (fino a 20 copie di DNA)
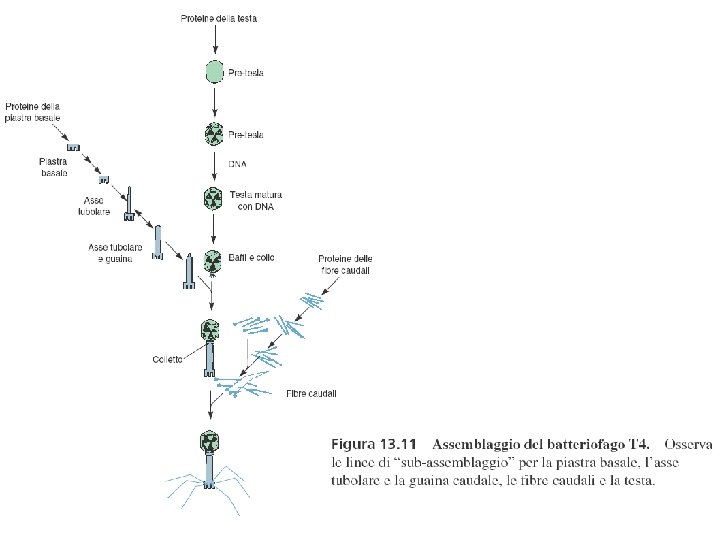
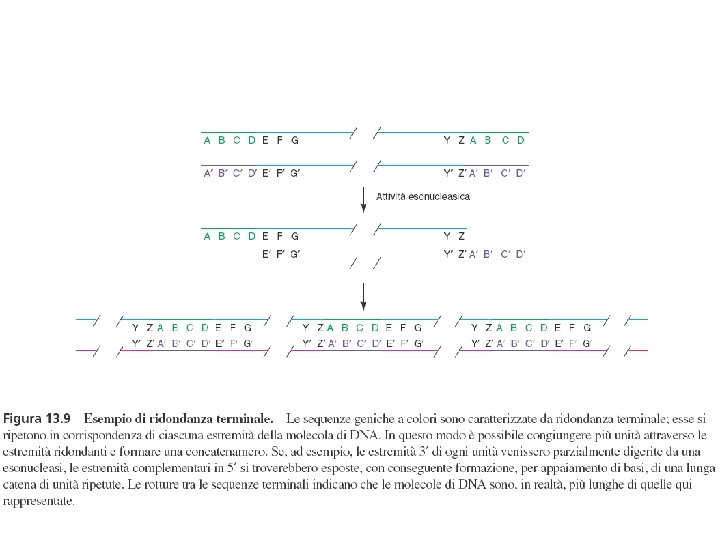
Assemblaggio delle particelle fagiche I geni tardivi dirigono la sintesi di tre tipi diversi di proteine: 1) proteine strutturali del fago 2) proteine che intervengono nell’assemblaggio ma non faranno parte del fago 3) proteine coinvolte nel processo di lisi batterica e liberazione dei fagi Le proteine necessarie all’assemblaggio vengono sintetizzate contemporameamente e sono utilizzate in “catene di montaggio” quasi indipendenti fino ad avere il fago maturo L’impacchettamento del DNA avviene quando la testa è matura: circa 500 mm di DNA devono essere inseriti in una cavità di appena 0, 1 mm. L’impacchettamento avviene secondo il meccanismo detto “a testa piena” a partire dai concatenameri sintetizzati. Quando il volume della testa è totalmente riempito il concatenamero viene tagliato. In questo modo entra il 102% del genoma virale. Questo processo è responsabile della ridondanza terminale e della permutazione circolare del genoma di T 4
Ridondanza terminale e permutazione circolare del genoma di T 4 123456789123456789123456789 102% 1234567891 102% 2345678912 102% 3456789123 102% 4567891234
X 174 e il fenomeno dei geni sovrapposti X 174 è un piccolo batteriofago icosaedrico con genoma a DNA circolare a singolo filamento È un esempio di come virus contenenti genomi di lunghezza limitata sfruttino al massimo l’informazione genetica attraverso il fenomeno dei geni sovrapposti I 5386 nucleotidi del genoma di X 174 non sarebbero sufficienti a codificare tutte le proteine virali identificate. Si è scoperto, quindi, che una sequenza può codificare anche per due proteine: i geni sono sovrapposti. 5386 Il gene E è contenuto all’interno di D su un altro schema di lettura. La proteina A* è parte della proteina A ed è ottenuta tramite un re-inizio della traduzione. A sintesi della forma replicativa A* inibizione della sintesi di DNA dell’ospite E lisi della cellula ospite
Il genoma di T 4 presenta ridondanza terminale
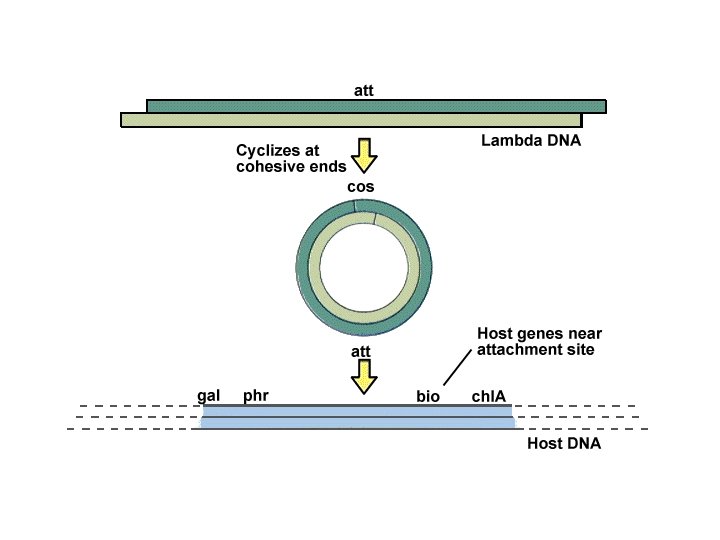
Batteriofagi temperati e lisogenia ciclo litico ciclo lisogenico induzione profago batterio lisogeno
Meccanismo di infezione dei batteriofagi (fagi) temperati se alcuni geni del DNA virale vengono espressi e ciò modifica il fenotipo cellulare si parla di CONVERSIONE LISOGENICA
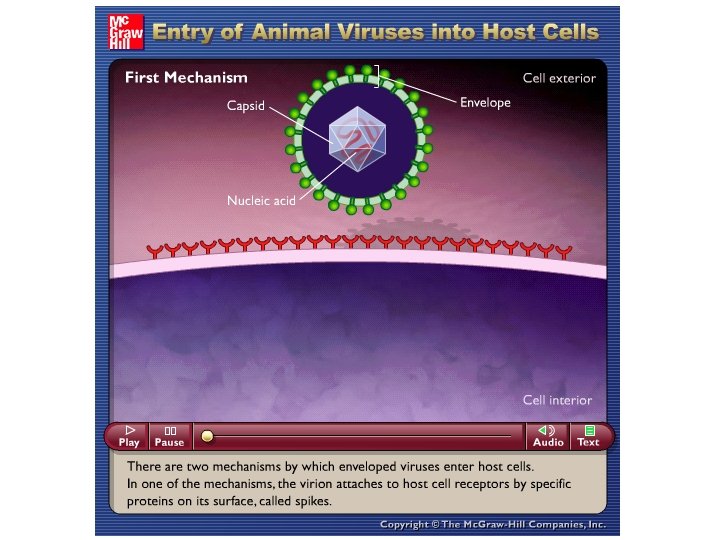
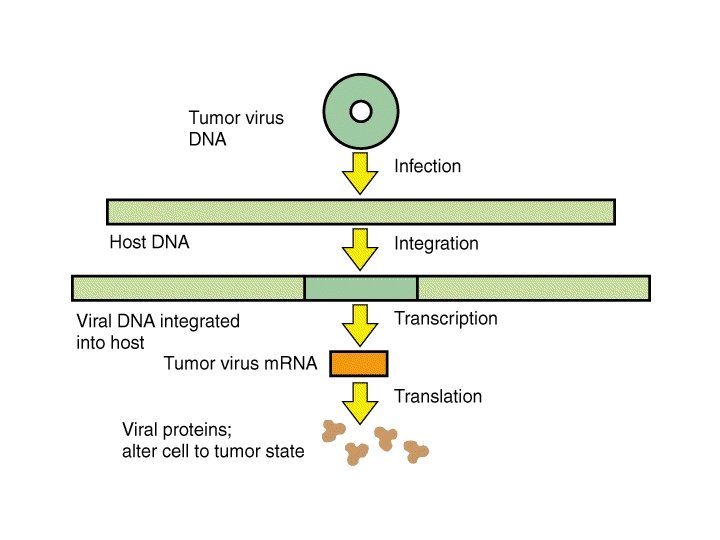
Meccanismo di infezione dei virus eucariotici
Infezioni acute: sono caratterizzate da una comparsa abbastanza rapida e da una durata ridotta; le cellule infettate generalmente vanno incontro a morte (effetto citopatico) e vengono liberati numerosi virioni. Le infezioni acute sono tipiche dei picornavirus, herpesvirus e adenovirus. Effetti citopatici 1) inibizione della sintesi di DNA, RNA e proteine 2) danneggiamento dei lisosomi con liberazione di enzimi idrolitici 3) alterazione della membrana plasmatica per inserimento delle proteine virali (attacco del sistema immunitario) 4) alta concentrazione delle proteine virali 5) rotture cromosomiche
Infezioni persistenti o croniche: possono durare anche molti anni. I virus possono replicarsi più lentamente senza causare una sintomatologia clinica. Sono presenti anticorpi diretti contro il virus infettante. Il virus dell’epatite B, il virus di Epstein-Barr e il virus del morbillo possono dare infezioni persistenti. Infezioni latenti: dovute a virus che, dopo una iniziale fase moltiplicativa, arrestano il proprio ciclo di sviluppo e diventano quiescenti. La fase di latenza può durare un certo periodo dopo il quale può verificarsi una recudescenza dell’infezione con una manifestazione spesso diversa da quella iniziale. I virus erpetici possono dare infezioni latenti. L’ herpes simplex di tipo 1 infetta in genere i bambini e rimane quiescente per anni a livello dei gangli del sistema nervoso; periodicamente può attivarsi e dare le caratteristiche lesioni erpetiche (herpes labiale) Il virus della varicella zoster (herpes zoster) provoca la varicella come prima manifestazione; non viene eliminato dal sistema immunitario ma diventa quiescente nei gangli dei nervi spinali. La manifesatzione ricorrente è il “fuoco di Sant’Antonio” per la riattivazione del virus a livello dei nervi intercostali
I virus responsabili dell'infezione appartengono alla famiglia degli orthomixovirus e si distinguono da quelli di altri disturbi infettivi per due caratteristiche. Innanzitutto perché non esiste un unico tipo di virus, ma ve ne sono di diversi, A, B e C: i primi due sono responsabili della classica forma di influenza, mentre il tipo C, generalmente asintomatico, provoca un'infezione simile al raffreddore. I virus di tipo A circolano sia nell'uomo che in altre specie animali (uccelli, maiali, cavalli) e sono a loro volta suddivisi in sottotipi. Di solito il virus si trasmette dagli uccelli al maiale e da quest'ultimo all'uomo. I virus di tipo B sono presenti solo nell'uomo e non esistono sottotipi distinti. In secondo luogo, i virus dell'influenza sono mutanti, cioè si modificano di anno in anno, obbligando così il nostro sistema immunitario ogni volta a produrre nuovi anticorpi in grado di affrontarli e sconfiggerli. Se ad esempio una persona ha superato un'infezione da virus di tipo A l'anno precedente, non è sicura di essere immune da ricadute l'anno successivo: lo stesso virus si può presentare modificato e l'organismo, per neutralizzarlo, deve produrre nuovi anticorpi. I virus dell'influenza sopravvivono solo nelle cellule delle prime vie respiratorie: naso, faringe e laringe. Ciò significa che se durante o in seguito ad un'influenza si sviluppa la broncopolmonite, i responsabili non possono essere i virus dell'influenza: in questo caso la malattia è dovuta ai batteri che, approfittando dello stato di debolezza della persona, ne attaccano i bronchi e i polmoni. Il virus che infetta volatili selvatici e domestici (tra cui i polli) si chiama "virus dell'influenza aviaria": di solito non infetta gli uomini, anche se, nel 1997 si sono verificati ad Hong Kong alcuni casi sporadici di influenza aviaria in persone che avevano avuto un contatto diretto con animali infetti (allevatori, macellatori e veterinari). Il virus dell'influenza aviaria, comunque, non si trasmette attraverso l'ingestione di carni o uova infette. Tipi di epidemia | Un po' di storia | Trasmissione. Tipi di epidemia Una caratteristica dell'influenza �la tendenza dei virus a modificare continuamente nel tempo le proprie caratteristiche: se le modifiche sono profonde in determinate circostanze ci possono essere importanti conseguenze per la popolazione che, non avendo mai incontrato il nuovo virus, �scarsamente immunizzata e si ammala pi�facilmente. Questo fenomeno pu�coincidere con la comparsa in tutti i gruppi di et�di grandi epidemie a livello mondiale, chiamate "pandemie". Le pandemie si verificano ad intervalli di tempo imprevedibili e in questo secolo sono avvenute nel 1918 (Spagnola, sottotipo H 1 N 1)), nel 1957 (Asiatica, sottotipo H 2 N 2) e nel 1968 (Hong Kong, sottotipo H 3 N 2). La pi�severa, la Spagnola, ha provocato almeno 20 milioni di morti. La comparsa di un ceppo di virus con proteine di superficie radicalmente nuove non �certo sufficiente per dire che si �verificata una pandemia. Occorre anche il nuovo virus sia capace di trasmettersi da uomo in modo efficace.
Sono conosciuti 3 tipi di virus influenzali: A = infettano molte specie animali B = infettano solo gli uomini C = asintomatici Hanno tutti 2 tipi di glicoproteine di superficie: emagglutinina (80% del totale) responsabile dell’interazione con la cellula ospite; neuramidasi coinvolta nel rilascio del virus dalla cellula infettata Il genoma è costituito da 8 pezzi di RNA a singolo filamento (il tipo C ha solo 7 segmenti di RNA). Il genoma è legato a proteine e ci sono 3 polimerasi per ogni segmento di RNA. Nuovi sottotipi e ceppi di virus influenzali emergono a causa del graduale accumularsi di mutazioni nelle regioni del virus che vengono riconosciute dal sistema immunitario dell’organismo ospite. Questo fenomeno prende il nome di deriva antigenica. Nei virus di tipo A può avvenire un altro tipo di fenomeno che accade quando 2 virus infettano la stessa cellula. In questo caso si può avere un rimescolamento dei segmenti che formano il genoma virale, causando una modifica profonda ed improvvisa del virus.