Prosm vypnte mobiln telefony Fyziologie rostlin Zkladn kurs

Prosím, vypněte mobilní telefony
")
Fyziologie rostlin Základní kurs 7 Transport Ing. Zuzana Balounová, Ph. D. (baloun@zf. jcu. cz)

MECHANISMUS PŘÍJMU A TRANSPORTU LÁTEK V ROSTLINĚ rozhraní mezi vnějším a vnitřním prostředím: toto rozhraní není vždy totožné s morfologickým povrchem rostliny - jejich orgánů: základní bariérou (o) a vnitřním (i) systémem je plazmalemma Přes ni vede primární vstup do buněčného prostředí. transport na krátké (vstup do buňky) střední dlouhé vzdálenosti.

Transport je vlastním procesem přemísťování látek a energie Přechod látky přes jednotkovou plochu rozhraní, daný množstvím převedené látky za čas označujeme jako tok a platí pro něj vztah: J=L. X L. . . vodivost prostředí pro transportovanou látku, X. . . příslušná hnací síla.
 a) se na její tvorbě podílí např. kinetická energie tepelného")
hnací síla transportu (X) a) se na její tvorbě podílí např. kinetická energie tepelného pohybu molekul, výpar gravitační síly vzlínavosti v kapilárních prostorách a ty formy pohybu prostředí, pro něž není hnací silou metabolický proces V těchto případech mluvíme o dějích pasivních. b) naproti tomu případy, kde X je produktem metabolických procesů jsou děje považovány za aktivní Tyto procesy mají svůj původ v samotném buněčném metabolismu, který vytváří energetické i látkové nerovnováhy (gradienty koncentrací), a ty jsou pak vlastní hnací silou aktivních transportních procesů Snížení metabolické aktivity (např. aplikací inhibitorů, snížení teploty aj. ) vede k poruše transportních procesů (přímo na membráně nebo nepřímo porušením aktivního konvekčního toku)

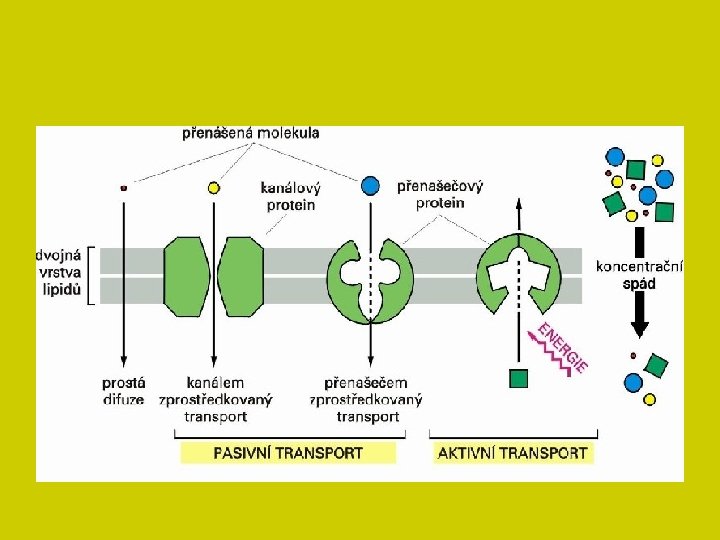
Pasivním transportem je především difúze Zdrojem energie je tepelný pohyb molekul (při stejné energii odpovídá rychlost pohybu molekul různé hmotnosti) směr pohybu má statický charakter a je určen rozdílem koncentrací látky na obou stranách rozhraní - koncentračním spádem cs = (co - ci). l, kde l. . . tloušťka vrstvy rozhraní Koncentrační spád se postupně v čase difúzí ruší, pokud jiné síly (např. metabolismus) znovu nevytvářejí koncentrační nerovnováhy. Procházejí-li ionty strukturami svázanými náboji (většinou zápornými), pak je transport kationtů urychlován, aniontů brzděn. Ionty opačných znamének však nejsou nikdy transportovány nezávisle, ale v roztoku se elektricky vyvažují.

Pohyb hydrofilních a iontově aktivních látek přes plazmalemmu umožňuje také usnadněná difúze: látka snadno pronikající lipidní dvojvrstvou membrány se na jedné straně (vnější) spojí s přenášeným iontem a vytvoří komplexní strukturu chelátu. Z něj se uvnitř zase uvolní výměnou za protony nebo hydroxylové ionty. Oba difúzní procesy samy o sobě jsou nezávislé na metabolické energii nebo na jiných zdrojích energie buňky. K typu pasivního transportu možno řadit i transport látek konvekčním tokem (pohybem) prostředí, nezávisle na tom, jakými silami je konvekční tok způsoben. pohybem prostředí není ovlivněn vlastní difúzní pohyb látky.

Formy pasivního transportu

Aktivní transport využívá speciálních iontově selektivních přenašečů za spotřeby makroergické vazby (ATP ev. jiné formy) Takový děj nabývá nutně povahy spřažené enzymatické reakce, jejíž kinetika vyhovuje enzymatické reakci prvého řádu (je-li ATP v nadbytku): S ohledem na současný (protisměrný) tok H+ a jejich vylučování mohou být tyto přenašeče aktivovány činností protonové pumpy.
 tok; elektrická kompenzace")
základní typy transportních systémů na rostlinných membránách uniport - samostatný (jednosměrný) tok; elektrická kompenzace může být zprostředkována třeba pasivním tokem dalších iontů; symport - spojený (jednosměrný) aktivní tok dvou iontů či iontů ve spojení s organickými látkami; antiport - spojený (protisměrný) aktivní tok dvou iontů souhlasného znaménka obrácená obdoba symportu (Na+/K+ - ATPáza, K+/H+ - ATPáza aj. ); elektroneutrální tok - přemísťují se molekuly bez náboje např. sacharóza Tím se dostávají neutrální látky do ionizovaného stavu, pokud se váží s elektricky nabitým iontem Takto se mohou transportovat přes membránu. Pro uniport H+ je popisován velmi účinný systém, tzv. protonová pumpa, která může aktivovat řadu dalších toků.

Transport může být popisován jak z vnějšku dovnitř tak zevnitř ven Transport protonů H+ běží z vnitřního prostředí do vnějšího proti němu jde transport ostatních kationtů (K+, Ca 2+ aj. ) dokud nedojde k rovnováze. Rovnováhy jsou dvojího typu: elektrické a koncentrační. Obě se vyrovnávají pasivním pohybem. Jsou však i nerovnováhy mezi vnějším a vnitřním prostředím, které se vyrovnávají aktivním procesem.
 Přenos")
1 - protonová pumpa, 2 - uniport, 3 - spřažený transport (symport, antiport) Přenos způsoben přenašečem
 - po koncentračním spádu. plyny (procházejí snadno")
Průchod látek membránou Prostou difúzí (buňka nereguluje) - po koncentračním spádu. plyny (procházejí snadno -důležité pro dýchání!) molekuly lipofilní povahy (uhlovodíky, např. benzen, mastné kyseliny, organické kyseliny) mezerami mezi bílkovinami a střední vrstvou bez ohledu na velikost molekul Čím méně rozpustné v tucích, tím pomaleji difundují membránou malé neutrální hydrofilní molekuly, které nejsou nabité ( voda, alkoholy, močovina, některé ionty a části aminokyselin, jednoduché cukry a nízkomolekulární bílkoviny ) jemné póry vyplněných vodou

Rychlost difúze - závisí na • koncentračním spádu látky na obou stranách membrány • teplotě • velikosti molekuly Difúze probíhá do vyrovnání koncentrace. D=K. 1/√M proto látky o malé molekulové hmotnosti difundují rychleji

Průchod látek membránou transmembránové kanály - specifické proteiny, umožňují průnik látek hydrofilních (např. iontové kanály pro Na+ a K+ ) • na průchod není nutné dodávat energii • některé jsou otevřené stále, jiné jen občas • jejich funkci lze regulovat.

Průchod látek membránou Elektrolyty - při pronikání ovlivněny elektrickými náboji pórů membrány: - vystlány buď + nebo - funkčními skupinami bílkovinných vrstev - ze svého okolí přitahují opačně nabité ionty a tak urychlují jejich pronikání membránou - v membráně převažují + , proto anionty do buňky snadněji než kationty - schopnost iontů pronikat plazmatickou membránou klesá se stupněm ionizace - slabé kyseliny a zásady pronikají lépe než silné - jednomocné kationty pronikají snadněji než vícemocné - u prvků stejného mocenství je jejich prostupnost membránovými póry omezena velikostí hydratovaných iontů Snadnost v pronikání (postupně klesá): NH 4+ K+ Na+ Li+ Mg 2+ Ba 2+ Sr 2+ Ca 2+ CNS- I- NO 3 CL-
")
Transport přes membránu prostou difúzí nepolární látky procházejí volně přes CPM, polární, hydrofilní (ionty) prostřednictvím iontových kanálů
 dochází k dočasnému spojení přenášené látky s tzv. \"nosičem„")
Usnadněnou difúzí (pasívní přenašečový transport) dochází k dočasnému spojení přenášené látky s tzv. "nosičem„ Komplex je rozpustný v lipoidní vrstvě membrány a šíří se jí po koncentračním spádu bez potřeby energie Tento proces je vysoce specifický Přenos aminokyselin, monosacharidů, disacharidů, fosfátových iontů někdy specifický, jindy nespecifický Transmembránová bílkovina váže přenášenou látku a jakýmsi způsobem ji dostane přes membránu. Snad to souvisí se změnou terciární nebo kvartérní struktury proteinu.

Aktivní transport vysvětluje nerovnoměrné rozložení látek uvnitř a vně buněčné membrány. . Přenašeči - enzymy zachytí molekulu vně a přenesou ji i proti koncentračnímu spádu dovnitř Tento proces vyžaduje energii, a tedy spotřebovává ATP syntetizovaný v mitochondriích vysoce selektivní proces, který je omezen na určité úseky membrány proces, kdy jsou přenášeny přes membrány látky proti koncentračnímu spádu za účasti transmembránového proteinu a za spotřeby energie proces působící v opačném směru než jednoduchá či usnadněná difuze typy aktivního transportu: membránové pumpy endocytóza exocytóza.

Na vazbu přenášené látky z prostředí mimo membránu je třeba energie Nahoře – nižší koncentrace iontů žádaných uvnitř buňky, kde je jejich koncentrace vyšší (Podobně pracuje např. protonová pumpa v membránách lysozómů. Transportem protonů dovnitř váčku se okyselí vnitřní prostředí a aktivují trávící enzymy)
 pumpy (umožňují přesun protonů přes membrány Tím se buď vytvoří protonový gradient")
Protonové (H+) pumpy (umožňují přesun protonů přes membrány Tím se buď vytvoří protonový gradient sloužící k jako zdroj energie k jiným reakcím (mitochondrie), nebo se vnitřní prostředí okyselí a tím se aktivují příslušné enzymy (lysozomy). Sodíko-draslíková pumpa Má za cíl udržet potřebné koncentrace Na+ a K+ iontů uvnitř buňky a mimo buňku Běžně se Na dostává do buňky snadno a jeho koncentrace převyšuje potřebu, zatímco K zůstává spíše mimo buňku, avšak je zde potřebný. Proto běžně 2 K+ čerpány do buňky, 3 Na+ čerpány z buňky za spotřeby 1 ATP Změny koncentrace iontů v intra- a extracellulárním prostředí umožňují rovněž vznik elektrického potenciálu

Na-K pumpa: za spotřeby 1 ATP se přes membránu přenesou 3 ionty Na+ ven a 2 ionty K+ dovnitř buňky. (Potassium - K, inside - uvnitř, outside- venku)

Sekundární aktivní transport jde o spřažení dvou pochodů, kdy jeden potřebuje dodání energie a druhý těží z takto vniklého koncentračního gradientu (symport a antiport)

Sekundární aktivní transport 1. 2. 3. 4. 5. 6. Využívá podobné proteiny jako usnadněná difuze Spojuje pohyb několika různých molekul do jednoho cyklu Kotransport přenáší dvě nebo více molekul přes membránu stejným směrem Opačný transport (counter transport) přenáší molekuly v opačných směrech Gradient vzniklý transportem jedné molekuly může být příčinou pohybu jiné molekuly proti jejímu vlastnímu gradientu Normální aktivní transport (Na-K ATPáza) vytváří silný gradient Na, který naopak umožňuje mnohé mechanismy sekundárního aktivního transportu (např. kotransport Na a glukózy)

Skupinová translokace jde o pochody, při nichž je transportovaný substrát chemicky změněn. Přenašeči jsou enzymy, které se na vnější straně s přenášenou látkou váží a tuto pak na vnitřní straně uvolňují jako produkt chemické reakce Energie na proces se získává z této chemické reakce
 endocytózy - dovnitř fagocytóza")
Obecně cytózy (splývání CPM buněk a váčků dovnitř nebo ven) endocytózy - dovnitř fagocytóza - pohlcování velkých částic nebo i celých buněk pinocytóza - zachycování drobných kapének tekuté potravy buněčným povrchem a transport dovnitř v podobě pinocytózních váčků. Po asimilaci zbytky jsou vypuzeny exocytózou. endocytóza s účastí receptorů (adaptiny - urychlení koncentrace přenášené látky a klatriny soudržnost váčků),
 pevné částice (nestrávené")
2. exocytózy – látka se dostává mimo buňku roztoku (produkt buňky) pevné částice (nestrávené zbytky) příklad exocytózy (vlevo) a endocytózy (vpravo) umožněných splýváním membrán.

Obrázek ukazuje soustavu tří polarizovaných buněk epitelu Pinocytózní váčky vznikající na jedné straně buňky vyloučí svůj obsah na druhé straně buňky.

TRANSPORT NA KRÁTKÉ VZDÁLENOSTI Přes plazmalemmu se látky dostávají aktivně nebo pasivně (Pinocytózy - neselektivní, ale může být velmi výkonným - přechod látek ze sousedních buněk systémem plazmodezmů (symplastická cesta). -minerální složky rostlinného těla nebo ty, které v minerální formě do buňky vstupují (zpravidla v iontové formě) -řada látek do buňky difunduje - tedy vstupuje pasivně. I v tomto případě může být v cestě difúzi nějaký filtrační systém (iontově selektivní kanály), který vede k určité „selektivitě“ - ovšem dokud se neanuluje hnací síla pro všechny složky tohoto systému, pak musí být výsledkem vždy rovnováha - pasivně vstupují i látky, které procházejí lipidní vrstvou membrány. Transport na krátké vzdálenosti v sobě zahrnuje základní mechanismus sycení symplastu, tj. vnitřního cytoplamatického systému z vnějšího prostoru, a proto celkový tok je závislý na povrchu protoplastu.

Vakuola (tonoplast - ten je sídlem aniontových transportních systémů a systémů pro transport organických látek. Tonoplast je dobře permeabilní pro kationty (zvláště K+), proto se vyrovnává nadbytek organických kyselin ve vakuole právě pasivní distribucí K+ iontu z cytoplazmatického prostoru. ve vakuole se hromadí ve zvýšeném množství jak anionty, tak kationty, které je potom doprovázejí, a tím celková koncentrace bude vyšší než v cytoplazmě a než ve vnějším prostředí. To vyvolává jevy, které vedou k osmotickým efektům v rostlinné buňce. Výsledkem toho je stav, že jak za kationty, tak za anionty se vyměňují iontové produkty vody (H+, OH- + CO 2 ---> HCO 3 -), které vyloučeny z buňky se opět vrací do nedisociované formy - vody. Z buňky ven se vylučuje sumárně voda a dovnitř vstupují soli. Soli mají osmotickou aktivitu, zatímco molekuly rozpouštědla ne. Vzniká tedy neustále přetlak osmoticky aktivních látek uvnitř rostlinné buňky, a tím dochází k intenzivnímu nasávání vody, zvětšuje se její objem a stoupá turgor To vyžaduje existenci pevné stěny buněčné s určitou elasticitou.

Membrány cytoplazmy rostlinné buňky nejsou během celého vývoje konstantní - mění se. V období objemového růstu se povrch buňky zvětšuje, zvětšuje se plazmalemma, vakuola a tonoplast. Právě díky hromadění K+ iontu v cytoplazmě a ve vakuole dochází k permanentnímu zvyšování turgoru v buňce, který je pracovním mechanismem při zvětšování jejich objemu Přirozený buněčný turgor je současně základem prostorového rozmístění všech bylinných orgánů rostliny turgescence - stav rostliny s dostatkem vody, solí atp.
 ve vnitřním vakuolárním roztoku není výsledkem postupného hromadění, ale")
Dosažení určité koncentrace látek (iontů) ve vnitřním vakuolárním roztoku není výsledkem postupného hromadění, ale dynamickou rovnováhou mezi tokem do buňky a výtokem do vnějšího prostředí Její úroveň se stabilizuje pro každý iont na jiné hodnotě. Některé ionty, např. vápenaté, mohou membránu stabilizovat, zvyšovat její semipermeabilitu ionty těžkých kovů mohou membránu narušovat a zvyšovat její permeabilitu (např. K+ v koncentraci nad 10 -3 M)

Existence membrány je tedy podmínkou, aby mohly vynikat rozdílné koncentrace iontů mezi vnějším a vnitřním prostředím, ale současně musí být tento efekt energeticky zajištěn. Na něm se podílí energetický metabolismus (z respiračních zdrojů nebo z fotosyntézy). energie může být využita buď na aktivní transport (na práci přenašeče) nebo na syntézu transportérů (chelatizačních látek). Vůči všem transportovaným kationtům je obecně kompetitivní proton (H+) oproti aniontům opět OH- nebo HCO 3 Pokud jde o aktivní transport iontů, potom zpravidla převládá transport kationtů (především K+)
 K")
Rozdíl elektrostatických aniontů je doplňován z organických kyselin (tvorbou kyseliny jablečné, kyseliny citronové) K tomu slouží fosfoenolpyruvátkinázá, případně další enzymy vázané v tonoplastu Tyto procesy vedou k biologickému pufrování cytoplazmy Jejím úkolem je udržovat p. H na neutrální hodnotě nezávisle na p. H okolních kompartmentů regulace p. H v buňce funguje na principu „biochemického p. H - statu“ Zvýšení p. H v cytoplazmě (> produkcí OH- při redukci NO 3 -) je způsobené syntézou silných kyselin (malát) z fosfoenolpyruvátu (PEP) za účasti PEPkarboxylázy Snížením p. H cytoplazmy (produkcí H+) se aktivuje malátový enzym, který dekarboxyluje malát a do prostředí kořene se uvolňuje CO 2 a OH-. Dekarboxylací malátu vznikne pyruvát a z něho neutrální produkt Optimum cytoplazmatického p. H (elektroneutralita) se tedy udržuje v mezích činností dvou enzymů (PEP-karboxyláza a malátový enzym).

Příjem a transport iontů v kořeni je selektivní proces. Kořeny rostlin se musí přizpůsobit různým exogenním koncentracím iontů tak, aby byla zabezpečena vhodná endogenní koncentrace i vzájemný poměr iontů v buňkách. V kořeni rostlin tedy probíhá intenzívní metabolismus, který je nutný k resorpci iontů. Vzhledem k tomu, že ve vakuolách kořenových buněk je koncentrace iontů vyšší než koncentrace iontů v tekuté půdní fázi, musí probíhat příjem živin proti koncentračnímu spádu, což je spojeno se spotřebou energie. Kořenové buňky musí mít k dispozici selektivní mechanismus příjmu živin, neboť jsou odkázány u celé řady živin na jejich velmi nízké koncentrace v půdním roztoku a na druhé straně je jim nabízena i relativně vysoká koncentrace prvků, které rostlina nezbytně potřebuje. Pro každou živinu je zapotřebí, aby byla v půdním roztoku dosažena alespoň její mezní koncentrace, jinak by její příjem mohl být omezen tak, že by se na rostlině začaly objevovat příznaky jejího nedostatku. Mezní koncentrace jsou pro jednotlivé živiny rozdílné.

V buněčných membránách jsou dva typy bílkovinných přenašečů, které jsou pro každý iont specifické v závislosti • na nízkých koncentracích (< 1 mmol. l-1) tzv. systém I, • na vysokých koncentracích(> 1 mmol. l-1), tzv. systém II. v plazmalemmě při nízkých koncentracích iontů jsou v činnosti jen mechanismy systému I s vysokou afinitou k iontům. V systému II při vysokých koncentracích iontů v půdním roztoku je systém I odpojený, ionty se transportují difúzí přes plazmalemmu a nosič funguje v tonoplastu

Lokalizace mechanismu příjmu iontů Předpokládá se, že mechanismy jednoho systému působí při jedné a druhého systému při druhé membráně
 mezi syntézou organických kyselin a")
Snahou každé živé buňky je zachovat si poměr (stechiometrický) mezi syntézou organických kyselin a uvolňováním H+ z buňky na udržení její elektroneutrality. regulace p. H v buňce se přímo spojuje s metabolismem, při kterém se buď produkuje nebo spotřebovává H+. Metabolické produkty, které obsahují C, H, O a N mají vždy negativní náboj v důsledku převahy karboxylových skupin nad NH 3. Přebytek karboxylových skupin musí být vyrovnán anorganickými kationty, aby byl zachován princip elektroneutrality potřebný k tomu, že většina enzymů cytoplazmy vykazuje optimum aktivity p. H mezi 6 -7. Ve vakuole je p. H nižší a pohybuje se v rozmezí 5 -6. Citlivost cytoplazmatických enzymů je tak velká, že kontrola p. H uvnitř buňky je základní podmínkou metabolismu.

Při vstupu NO 3 - nebo SO 42 - se z buňky uvolňuje OHpři vstupu NH 4+ se vylučuje H+ Vtok NO 3 - se vybalancuje výtokem OHBěhem respirace buňky p. H cytoplazmy klesá v důsledku uvolňování H+, který se transportuje z mitochondrií do cytoplazmy, případně do vnějšího prostředí Současně se antiportem transportuje dovnitř K+, který zvyšuje turgor a cytoplazmatické p. H začne narůstat. Zvýšené p. H podmiňuje syntézu organických kyselin (malát), a tím i produkci H+ (COO-, H+) Výměnou za H+/K+ přijatý K může neutralizovat kyseliny (K-malát) nebo se ukládá do vakuoly. Syntéza organických kyselin probíhá potud, dokud se vyměňuje vytěsněný proton za K+ anebo než se produkty této syntézy převedou do vakuoly. Toto je pouze příklad možného propojení metabolismu buňky k p. H.

Změny p. H v buňce vedou k produkci H+ nebo OH-, které se uvolňují jako odezva na signály mající původ v metabolismu. Vlastností protonu H+ je schopnost recirkulace, tj. opětovně vstupovat do buňky podle jejího požadavku na neutralizaci volných OH- skupin Na rozdíl od H+ iontu OH- ionty, které se uvolňují z buňky při zvýšení cytoplazmatického p. H, nepodléhají recirkulaci a jsou tedy pro buňku ztracené. Ionty, které vstupují do buňky jsou postupně metabolicky utilizovány, různě zabudovávány a přitom se může měnit jejich iontový charakter (např. z aniontu NO 3 - se tvoří kationická aminová skupina atp. ) a stávají se členy systému cytoplazmy. Řada z nich může být podle potřeby znovu i opakovaně (několikrát) přeměňována a přitom mění svou funkci v organických látkách, ale i v živých systémech. Tyto procesy přeměny látek označujeme jako reutilizaci a jsou běžné při klíčení semen, rašení pupenů, vegetativním množení, ale i v průběhu ontogenetického vývoje rostlin.

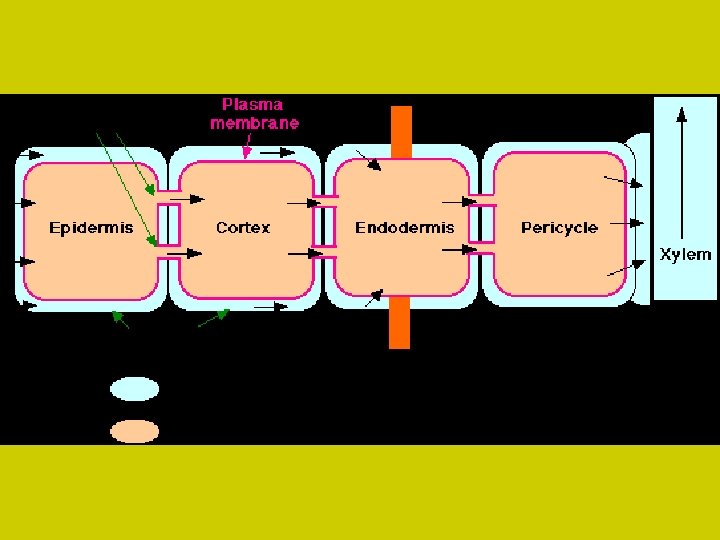
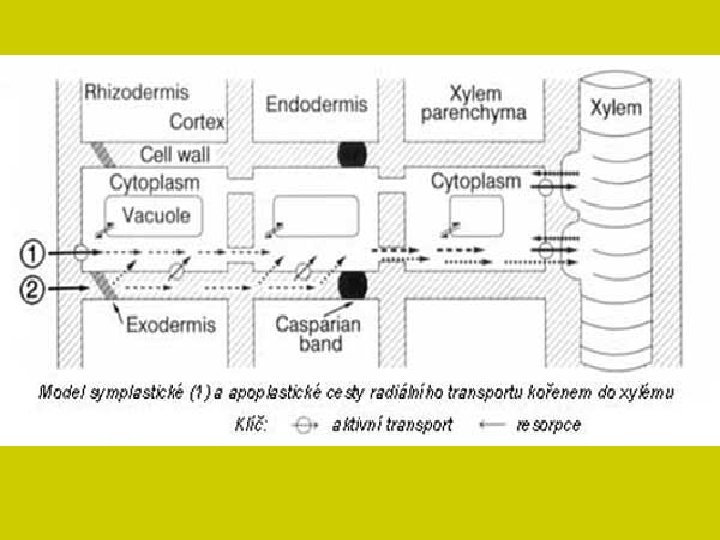
TRANSPORT NA STŘEDNÍ VZDÁLENOSTI Iont přijatý z vnějšího prostředí buňky může být • okamžitě zapojen (utilizován) v metabolismu • uložen ve vakuole • nebo transportován do dalších buněk. Transport na střední vzdálenosti u vícebuněčných rostlin zajišťuje transport přijatých iontů mezi vnitřním a vnějším prostorem dále dovnitř rostlinných pletiv. symplast a apoplast Oba umožňují pohyb iontů spojitým prostorem, avšak současně to jsou systémy, které ionty vyvazují a různě přetvářejí jejich původní iontové formy. Situace se mění podle stáří rostliny a typu diferenciace jejich pletiv. Pohyblivost iontu v obou systémech je omezená a rychlost je velmi malá.

symplast Iontový transport od buňky k buňce míjí vakuoly a postupuje podél souvislého řetězce živých protoplastů, které se stýkají plazmodesmami Tím je vytvořen z celého cytoplazmatického prostředí určitého orgánu vodivý systém. Kanálky tvořené plazmodesmami (vnější průměr 40 nm, vnitřní 25 nm) nejsou pro ionty za všech podmínek průchodné. Přesto vede tato cesta symplastem až do vodivé soustavy středního válce. Tam sítkovice a ostatní pomocné buňky přijímají ionty přímo dotykem se symplastem. Ionty jsou pasivně transportovány ve směru koncentračního spádu jednak do odumřelých cév a vedle toho ionty do cév aktivně vylučují parenchymatické buňky.
 do")
apoplazmatický transport prostorem stěn buněčných, které mohou komunikovat s vnějším prostředím (vodným roztokem) do různých vnitřních systémů, vzduchových (plynných) kanálů a průduchy do vnějšího prostředí Má tedy dvě složky: Jedna obsahuje plyny (intercelulární systém) druhá vodný roztok hydrofilních látek (stěny buněčné) Prvý systém bývá označován jako plynný (gas systém, GS), druhý jako volný prostor (pro transport vodných roztoků), tzv. „free space“ (FS).
, nýbrž dva podsystémy: především")
apoplazmatický transport V apoplazmatickém systému není jenom jeden prostor (FS), nýbrž dva podsystémy: především vlastní vodný prostor (WFS) dále tzv. Donnanův volný prostor (DFS), který na rozdíl od prvého nese fixované náboje, především záporného charakteru (karboxylové zbytky pektinových látek). DFS funguje jako iontoměničový systém a není strukturně oddělen od vodného volného prostoru. Volně pohybující se ionty mohou přecházet do Donnanova prostoru a vyměňovat se s ionty fixovanými na strukturách stěn buněčných. Pokud jsou tam na monovalentním aniontovaném zbytku vázány polyvalentní kationty, pak se mohou na kationty opět vázat jiné volné anionty. Tak vzniká vlastně přepolarizování iontoměničového systému stěn buněčných. Stoupá počet interakcí mezi ionty s různými znaménky a stěnou buněčnou.

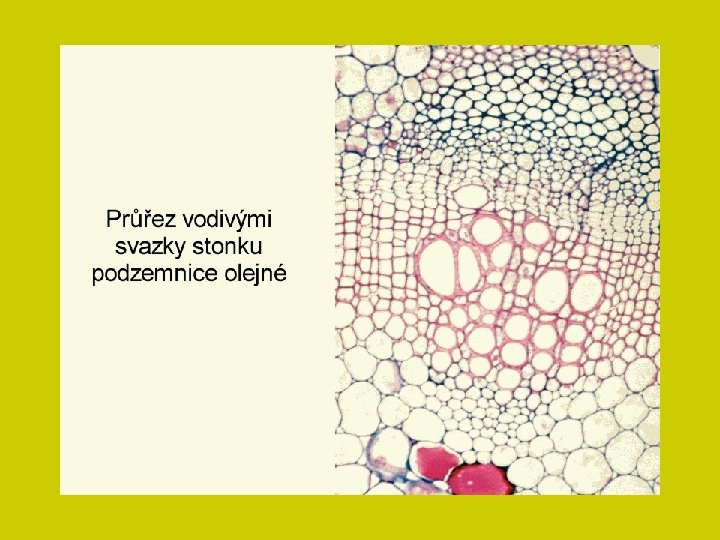
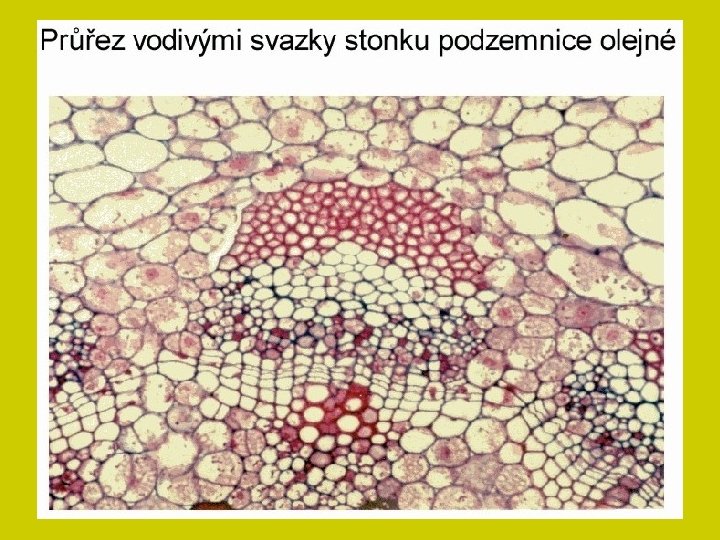
Ionty i soli se pohybují xylémem a transpirační proud je v rostlině rozvádí. Na konci cévních svazků difundují buněčnými stěnami k povrchům protoplastů ve svazkovém parenchymu a aktivním transportem přestupují do parenchymatických buněk. Transport živin mezi buňkami prochází tak opět symplastem a některé soli jsou přitom ukládány do vakuol.

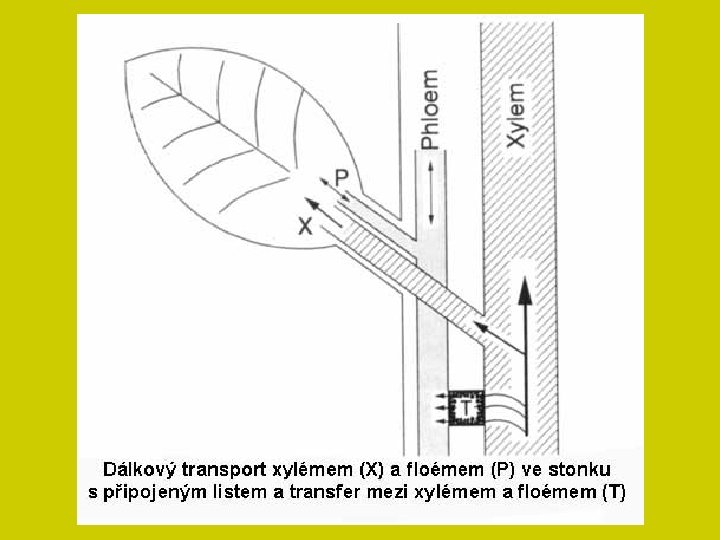
Floém Obě dálkové translokační soustavy jsou na mnoha místech propojeny zejména v kořenech a stonkových nodech. Spolu s tokem metabolitů jsou anorganické látky dodávány tam, kde je jich nejvíce třeba. Sítkovicemi se zprostředkuje zejména přerozdělení těch látek, které již jednou rostlina přijala. Tato redistribuce je pro různé látky rozdílně obtížná. Tak živiny vázané v organických sloučeninách (např. N, P, S) se mohou translokovat jako ionty alkalických kovů, Cl- a NO 3 -. Naopak obtížná je translokace těžkých kovů a iontů alkalických zemin, zejména vápníku. Proto se tyto prvky hromadí v listech, kde končí xylémová cesta pro translokaci živin.

DÁLKOVÝ TRANSPORT ionty i soli se pohybují xylémem a transpirační proud je v rostlině rozvádí Na konci cévních svazků difundují buněčnými stěnami k povrchům protoplastů ve svazkovém parenchymu a aktivním transportem přestupují do parenchymatických buněk. Transport živin mezi buňkami prochází tak opět symplastem a některé soli jsou přitom ukládány do vakuol.

DÁLKOVÝ TRANSPORT Vedle xylému je další významnou cestou floém. Obě dálkové translokační soustavy jsou na mnoha místech propojeny zejména v kořenech a stonkových nodech. Spolu s tokem metabolitů jsou anorganické látky dodávány tam, kde je jich nejvíce třeba. Sítkovicemi se zprostředkuje zejména přerozdělení těch látek, které již jednou rostlina přijala. Tato redistribuce je pro různé látky rozdílně obtížná. Tak živiny vázané v organických sloučeninách (např. N, P, S) se mohou translokovat jako ionty alkalických kovů, Cl- a NO 3 -. Naopak obtížná je translokace těžkých kovů a iontů alkalických zemin, zejména vápníku. Proto se tyto prvky hromadí v listech, kde končí xylémová cesta pro translokaci živin.
- Slides: 54