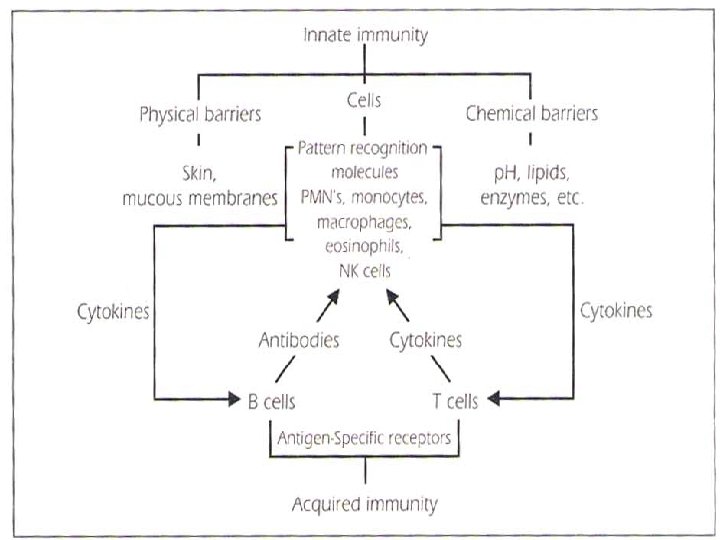
ADAPTIVE IMMUNITY ADAPTIVE ACQUIRED IMMUNITY CHARACTERISTICS Generation of

 IMMUNITY CHARACTERISTICS: Generation of antigen – specific lymphocytes “effector cells” Generation of")


 Cytotoxic or Killer (CD 8+) Memory Regulatory (suppressor)")
 They assist other white blood cells in immunologic processes,")
 Once activated, they divide rapidly and secrete small proteins")

 destroy")

 express CD 45 RO but lack expression")
 differ from the other memory subsets in that")
 Their major role is to shut down T")









 Fab VH VL CH 1 CL CH 2 Fc CH 3")
 J-Chain 10 Heavy and 10 Light chains 10 Fab and 5")
 Monomeric- Serum Ig. A ~15 -20% of serum immunoglobulins 2 Heavy")
 J-Chain Secretory Component 4 heavy chains and 4 light chains")
 CH 4 2 Heavy and 2 Light chains Mol. Wt. ~190,")
 2 heavy and 2 light chains <1% of serum immunoglobulins Serves")



























")








- Slides: 74
ADAPTIVE IMMUNITY
ADAPTIVE (ACQUIRED) IMMUNITY CHARACTERISTICS: Generation of antigen – specific lymphocytes “effector cells” Generation of Memory cells which prevent reinfection with the same organism. It takes more time to develop “>96 h” It exhibits specificity against foreign substances. Consists from cellular (lymphocytes) and humeral (antibodies) immunity
CELLULAR ADAPTIVE IMMUNITY: LYMPHOCYTES
T CELLS A T cell, or T lymphocyte, is a type of lymphocyte that plays a central role in cell-mediated immunity. T cells can be distinguished from other lymphocytes, such as B cells and natural killer cells, by the presence of a T-cell receptor on the cell surface. They are called T cells because they mature in the thymus from thymocytes (although some also mature in the tonsils). The several subsets of T cells each have a distinct function.
T CELL TYPES Helper (CD 4+) Cytotoxic or Killer (CD 8+) Memory Regulatory (suppressor) Natural killer T cell Mucosal associated invariant Gamma delta T cells
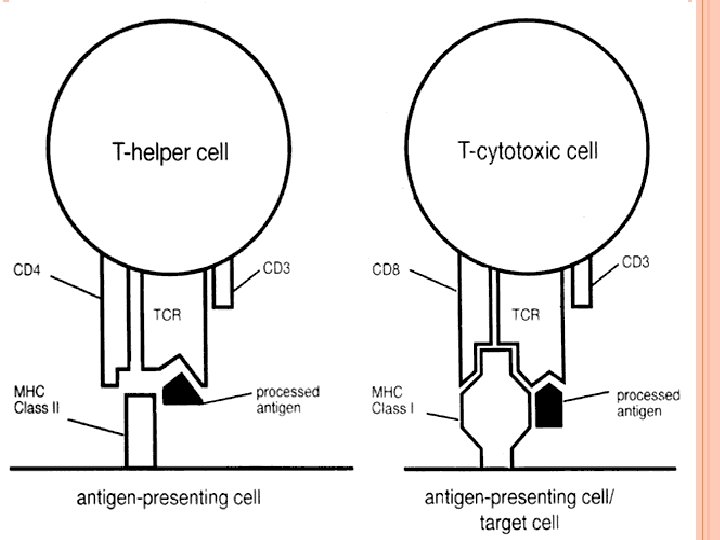
T HELPER CELLS (TH CELLS) They assist other white blood cells in immunologic processes, including maturation of B cells into plasma cells and memory B cells and activation of cytotoxic T cells and macrophages. These cells are also known as CD 4+ T cells because they express the CD 4 glycoprotein on their surfaces. Helper T cells become activated when they are presented with peptide antigens by MHC class II molecules, which are expressed on the surface of antigen-presenting cells (APCs).
T HELPER CELLS (TH CELLS) Once activated, they divide rapidly and secrete small proteins called cytokines that regulate or assist in the active immune response. These cells can differentiate into one of several subtypes, including TH 1, TH 2, TH 3, TH 17, TH 9, or TFH, which secrete different cytokines to facilitate different types of immune responses. Signaling from the APC directs T cells into particular subtypes
Cytokine secreted by APC Transcription factors • Intracellular pathogens • Macrophage activation • Extracellular pathogens • Antibodies production • • • Helminths and allergy Autoimmunity Tumor suppression Extracellular bacteria and fungi Neutrophils activation • promote tissue repair or remodeling. pathogenesis of inflammatory skin disorder • Long term humoral immunity • Inhibit dendritic cells • Suppress T cells • Prevent autoimmunity
T CYTOTOXIC Cytotoxic T cells (TC cells, CTLs, T-killer cells, killer T cells) destroy virus-infected cells and tumor cells, and are also implicated in transplant rejection. These cells are also known as CD 8+ T cells since they express the CD 8 glycoprotein at their surfaces. These cells recognize their targets by binding to antigen associated with MHC class I molecules, which are present on the surface of all nucleated cells. Through IL-10, adenosine, and other molecules secreted by regulatory T cells, the CD 8+ cells can be inactivated to an anergic state, which prevents autoimmune diseases.
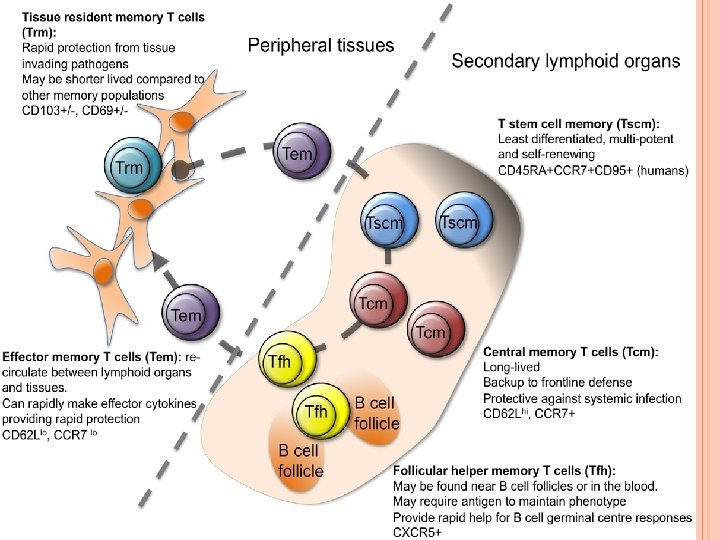
MEMORY T CELLS The single unifying theme for all memory T cell subtypes is that they are long-lived and can quickly expand to large numbers of effector T cells upon re-exposure to their cognate antigen. By this mechanism they provide the immune system with "memory" against previously encountered pathogens. Memory T cells may be either CD 4+ or CD 8+ and usually express CD 45 RO. Memory T cell common subtypes: 1. Central memory T cells (TCM cells) express CD 45 RO, C-C chemokine receptor type 7 (CCR 7), and L-selectin (CD 62 L). Central memory T cells also have intermediate to high expression of CD 44. This memory subpopulation is commonly found in the lymph nodes and in the peripheral circulation.
2. Effector memory T cells (TEM cells) express CD 45 RO but lack expression of CCR 7 and L-selectin. They also have intermediate to high expression of CD 44. These memory T cells lack lymph nodehoming receptors and are thus found in the peripheral circulation and tissues. 3. Tissue resident memory T cells (TRM) occupy tissues (skin, lung, gastrointestinal tract, etc. ) without recirculating. One cell surface marker that has been associated with TRM is the integrin αeβ 7. These cells are thought to play a major role in protective immunity against pathogens. Dysfunctional TRM cells have been implicated in autoimmune diseases, such as psoriasis, rheumatoid arthritis, inflammatory bowel disease.
4. Virtual memory T cells (TVM) differ from the other memory subsets in that they do not originate following a strong clonal expansion event. Thus, although this population as a whole is abundant within the peripheral circulation, individual virtual memory T cell clones reside at relatively low frequencies.
REGULATORY T CELLS (SUPPRESSOR T CELLS) Their major role is to shut down T cell-mediated immunity toward the end of an immune reaction and to suppress autoreactive T cells that escaped the process of negative selection in the thymus. Crucial for the maintenance of immunological tolerance. Regulatory T cells can develop either during normal development in the thymus, and are then known as thymic Treg cells, or can be induced peripherally and are called peripherally derived Treg cells. These two subsets were previously called "naturally occurring", and "adaptive" or "induced", respectively.
Both subsets require the expression of the transcription factor FOXP 3 which can be used to identify the cells. Mutations of the FOXP 3 gene can prevent regulatory T cell development, causing the fatal autoimmune disease (IPEX). Several other types of T cell have suppressive activity, but do not express FOXP 3. These include Tr 1 cells and Th 3 cells, which are thought to originate during an immune response and act by producing suppressive molecules. Tr 1 cells are associated with IL-10, and Th 3 cells are associated with TGFbeta. Recently, Treg 17 cells have been added to this list.
NATURAL KILLER T CELL Natural killer T cells bridge the adaptive immune system with the innate immune system. Unlike conventional T cells that recognize peptide antigens presented by major histocompatibility complex (MHC) molecules, NKT cells recognize glycolipid antigen presented by a molecule called CD 1 d.
NATURAL KILLER T CELL Once activated, these cells can perform functions ascribed to both Th and Tc cells (i. e. , cytokine production and release of cytolytic/cell killing molecules). They are also able to recognize and eliminate some tumor cells and cells infected with herpes viruses. (NKT cells – not to be confused with natural killer cells of the innate immune system which is CD 3 negative).
GAMMA DELTA T CELLS The majority of human T cells, termed alpha beta T cells (αβ T cells), rearrange their alpha and beta chains on the cell receptor and are part of the adaptive immune system. Specialized gamma delta T cells, (a small minority of T cells in the human body, more frequent in ruminants), have invariant T-cell receptors with limited diversity, that can effectively present antigens to other T cells and are considered to be part of the innate immune system. This group of T cells is much less common in humans and mice (about 2% of total T cells)
B CELLS also known as B lymphocytes, are a type of white blood cell of the lymphocyte subtype. They function in the humoral immunity component of the adaptive immune system by secreting antibodies. Additionally, B cells present antigen (they are also classified as professional antigen-presenting cells (APCs)) and secrete cytokines. In mammals, B cells mature in the bone marrow, which is at the core of most bones
B CELLS In birds, B cells mature in the bursa of Fabricius, a lymphoid organ. (The "B" from B cells comes from the name of this organ, where it was first discovered by Chang and Glick, and not from bone marrow as commonly believed). B cells, unlike the other two classes of lymphocytes, T cells and natural killer cells, express B cell receptors (BCRs) on their cell membrane. BCRs allow the B cell to bind to a specific antigen, against which it will initiate an antibody response. B cells have many subclasses. Brief info about them could be found in the slide notes.
IL-21 & others
HUMORAL ADAPTIVE IMMUNITY: ANTIBODIES
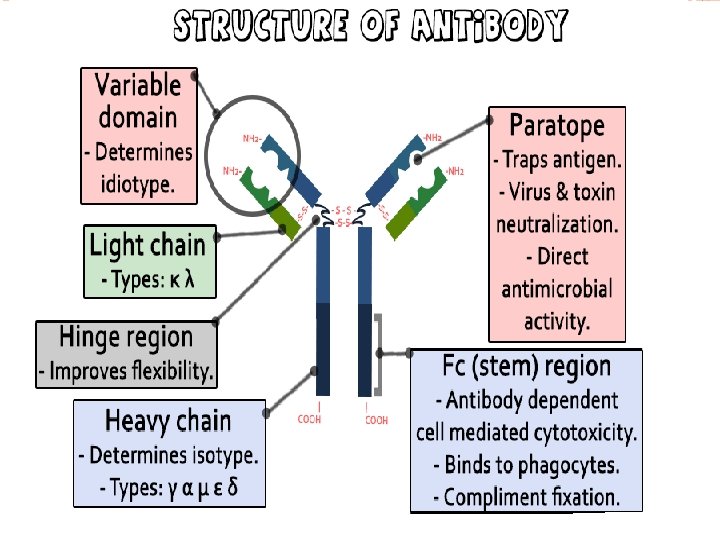
WHAT IS AN ANTIBODY? Product of adaptive immunity Made specifically to bind a unique antigenic epitope (also called an antigenic determinant) Possesses an antigen binding site A product of the Plasma cell Members of immunoglobulins the class of proteins called
IMMUNOGLOBULIN G (IGG) Fab VH VL CH 1 CL CH 2 Fc CH 3 2 Heavy and 2 Light chains 2 Fab and 1 Fc fragment 4 Subclasses (Ig. G 1, Ig. G 2, Ig. G 3, Ig. G 4) Mol. Wt. 150, 000 ~70 -75% of serum immunoglobulin. The major antibody of the secondary immune response Change in affinity with time
IMMUNOGLOBULIN M (IGM) J-Chain 10 Heavy and 10 Light chains 10 Fab and 5 Fc fragments Mol. Wt. ~900, 000 <10% of serum immunoglobulin. Single J Chain (15 k. DA) The predominant "early" antibody Most primitive immunoglobulin No change in affinity with time
IMMUNOGLOBULIN A (IGA) Monomeric- Serum Ig. A ~15 -20% of serum immunoglobulins 2 Heavy and 2 Light 2 Subclasses (Ig. A 1 and Ig. A 2) Found in serum
SECRETORY IMMUNOGLOBULIN A (SIGA) J-Chain Secretory Component 4 heavy chains and 4 light chains (dimeric) J-Chain and secretory component The major immunoglobulin of secretions Not found in serum
IMMUNOGLOBULIN E (IGE) CH 4 2 Heavy and 2 Light chains Mol. Wt. ~190, 000 Trace serum protein Note CH 4 region on H chain Associated with atopic or anaphylactic hypersensitivity May play role in immunity to helminthic parasites
IMMUNOGLOBULIN D (IGD) 2 heavy and 2 light chains <1% of serum immunoglobulins Serves as a membrane receptor on B lymphocytes May play role in antigenstimulated lymphocyte differentiation
Class Subclass Ig. A Ig. D Ig. E Ig. G Ig. M Description 2 Found in mucosal areas, such as the gut, respiratory tract and urogenital tract, and prevents colonization by pathogens. Also found in saliva, tears, and breast milk. 1 Functions mainly as an antigen receptor on B cells that have not been exposed to antigens. It has been shown to activate basophils and mast cells to produce antimicrobial factors. 1 Binds to allergens and triggers histamine release from mas cells and basophils, and is involved in allergy. Also protects against parasitic worms 4 In its four forms, provides most of the antibody-based immunity against invading pathogens. The only antibody capable of crossing the placenta to give passive immunity to the fetus. 1 Expressed on the surface of B cells (monomer) and in a secreted form (pentamer) with very high avidity. Eliminates pathogens in the early stages of B cell-mediated (humoral) immunity before there is sufficient Ig. G.
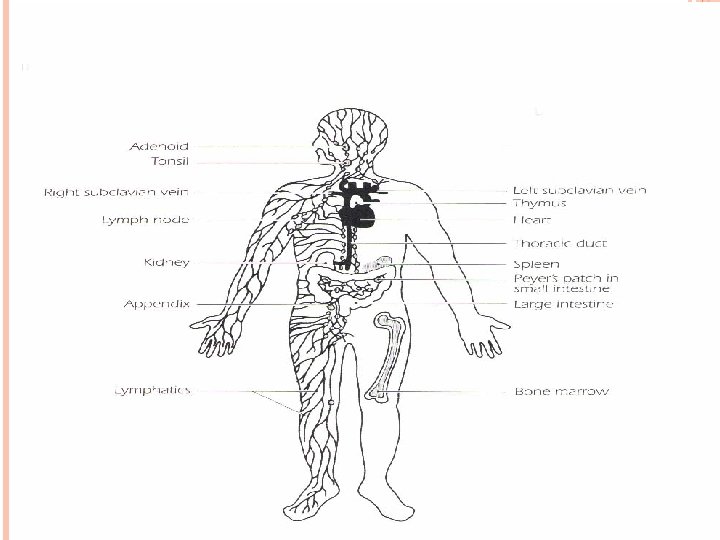
LYMPHATIC ORGANS
LYMPHATIC ORGANS These organs are the sites in which lymphocyte maturation, differentiation and proliferation take place. A. The primary “central” lymphoid organs. . . Thymus gland bone marrow. Maturation of T and B lymphocytes into antigen – recognizing lymphocytes and acquiring their antigen. B. • Secondary “peripheral” lymphoid organs They are those organs where antigen–driven proliferation and differentiation occur. These include lymph nodes, spleen, gut-associated lymphoid tissues such as tonsils.
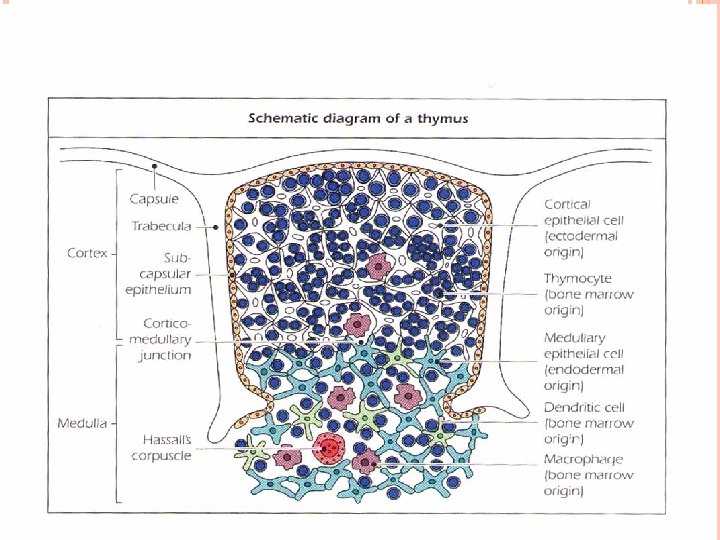
THYMUS GLAND Progenitor cells from the bone marrow migrate to the thymus, where they differentiate into T lymphocytes The thymus gland is a lymphoepitheloid bilobed structure It is derived from the endoderm of the third and fourth pharyngeal pouches. During fetal development, the size of the thymus increases, the growth continues until puberty, thereafter, the thymus undergoes atrophy with aging. The cortex and medulla are infiltrated with thymocytes. T lymphocytes mature in cortex and migrate to the medulla. Macrophages and dendritic cells are found in medulla.
Macrophages are involved in clearing apoptotic thymocytes. Mature T lymphocytes migrate to the secondary lymphoid organs in which they encounter and respond to foreign antigens. T lymphocytes recognize and respond to foreign antigen via their specific receptors “TCR” which is acquired during differentiation in the thymus. Only 5 -10% of maturing lymphocytes survive and eventually leave thymus, 90 -95% of all thymocytes die in the thymus. Lymphocytes which die have developed specificity to self structures or have failed to make functional receptors and, therefore, are eliminated.
BONE MARROW In embryonic life , B cells differentiate from hematopoietic stem cells in the fetal liver. After birth this function moves to the bone marrow. B cells mature in bone marrow and bear antigen – specific receptors that have a structure and specificity identical to the antibody later synthesize by that B cells. The mature B cells are transported by the circulating blood to the secondary lymphoid organs, where they encounter and respond to foreign antigens.
SECONDARY LYMPHOID ORGANS The major secondary lymphoid organs are the spleen and lymph nodes. In addition, tonsils, appendix, peyer’s patches, and lymphoid aggregates spread throughout mucosal tissue “mucosal – associated lymphoid tissue – MALT”. Those associated with the gut “gut – associated lymphoid tissue – GALT, bronchus – associated lymphoid tissue BALT” The secondary lymphoid organs have two major functions: 1. Trapping and concentrating foreign substances 2. The main sites of production of antibodies and the induction of antigen – specific T Lymphocytes.
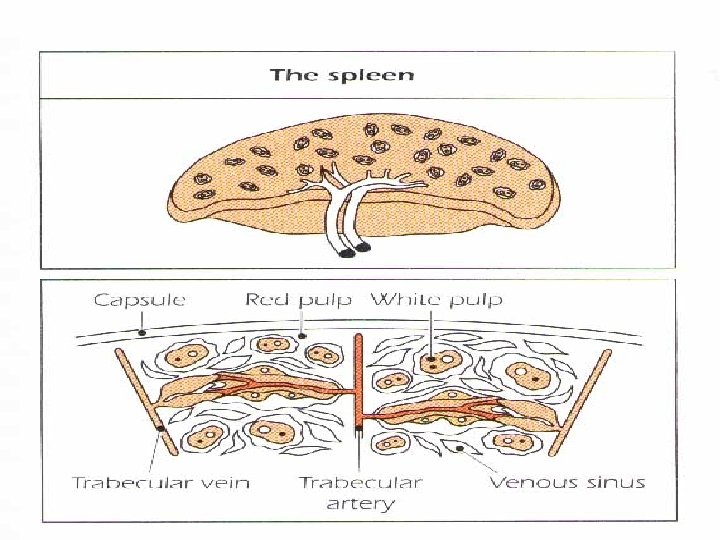
THE SPLEEN The largest organ of the secondary lymphoid organs. Traps and concentrates foreign substances carried in the blood. The major organ of antibody synthesis White pulp is rich in lymphoid cells Red pulp contains many sinuses, large quantities of RBCs and macrophages , some lymphocytes.
THE SPLEEN B – cells are present mainly in germinal centers, T cells in the peripheral region. Approximately 50% of spleen cells are B lymphocytes, 30 -40% are T lymphocytes. After antigenic stimulation, the germinal centers contain large numbers of B cells and plasma cells, these cells synthesize and release antibodies.
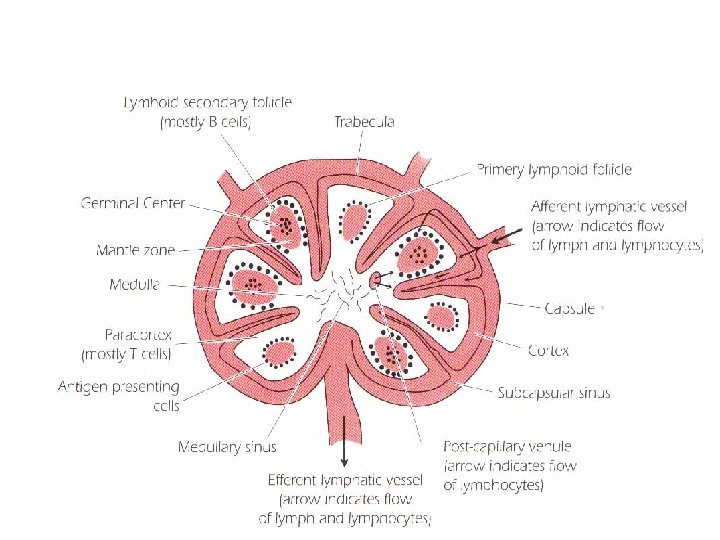
LYMPH NODES lymph nodes are small, ovoid structures They are close to major junction of the lymphatic channels, which are connected to the thoracic duct. The thoracic duct transports lymph and lymphocytes to the vena cava, the vessel that carries blood to the right side of the heart, from where it is redistributed throughout the body. The cortical region contains primary lymphoid follicles.
LYMPH NODES After antigenic stimulation, they enlarge to form secondary lymphoid follicles with germinal centers containing B lymphocytes. They generate clones of cells with higher affinity receptors “antibody” for the antigenic epitope that triggered the initial response. The deep cortical area or Para cortical region contains T cells and dendritic cells which present antigen fragments to T cells The medullary area of the lymph node contains antibody – secreting plasma cells that have travelled from the cortex to the medulla via lymphatic vessels.
LYMPHOCYTE CIRCULATION
The migration of lymphocytes between various lymphoid and nonlymphoid tissue and their homing to a particular site is highly regulated by means of various cell – surface adhesion molecules “CAMs” and receptors to these molecules. Blood lymphocytes cross the endothelial vascular lining of post capillary vascular sites termed high endothelial venules “HEVs” this process is called extravasation. Recirculating lymphocytes selectively bind to specific receptors on the HEV of lymphoid tissue. Recirculating monocytes and granulocytes also express adhesion molecule receptors and migrate to tissue sites using a similar mechanism.
The traffic of lymphocytes between lymphoid and non lymphoid tissue ensures that on exposure to an antigen, the antigen and the lymphocytes specific to that antigen are sequestered in the lymphoid tissue, where the lymphocytes undergo proliferation, and differentiation. The differentiated cells (memory cells) leave the lymphoid organ and are dessiminated in the body to reconcentrate at the site where antigen persists and, at that place, exert their protective function.
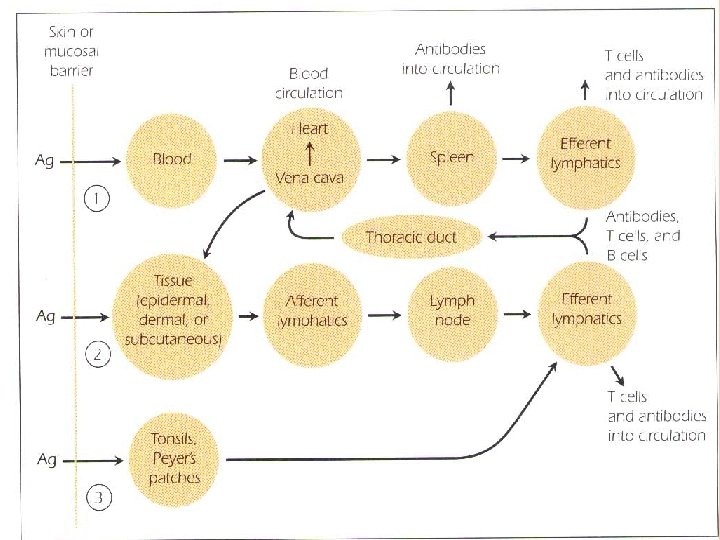
THE FATE OF ANTIGEN AFTER PENETRATION The RES traps foreign antigens and subject them to ingestion and degradation by the phagocytic cells of the system There is constant movement of lymphocytes which direct them to be deposited in strategic places along the lymphatic vessels. The system also provides Loci “ the secondary lymphoid organs” where antigen, macrophages, T cells and B cells can interact within a very small area to initiate an immune response.
Three major route may be followed by an antigen after it has penetrated the interior of the body: 1. If the antigen enters the body through the blood stream, it is carried to spleen, where it interacts with APCs “dendritic cells, macrophages , B cells”. APCs activate – antigen – specific T cells. The spleen releases antibodies into the circulation. Lymphocytes leave the spleen through the efferent lymphatics to reenter the circulation via the thoracic duct. 2. The antigen may lodge in epidermal , dermal or subcutaneons tissue, where it may cause an inflammatory response.
3. When antigen is trapped by APCs, it is transported through the afferent lymphatic channels into the regional lymph nodes. In the lymph node, the antigen, macrophages, dendritic cells, T cells and B cells interact to generate an immune response. Antigen –specific T cells and antibodies which have been synthesised in the lymph node , enter the circulation via the thoracic duct. Antitgen may enter GIT or respiratory tract, where it lodges in the MALT. Where it interact with macrophages and lymphocytes. Antibodies are deposited in local tissue, in addition, lymphocytes entering the efferent lymphatics are carried through the thoracic duct to the circulation and redistributed to various tissues.
ANTIGENS
IMMUNOGENS AND ANTIGENS The compound that evokes the response is referred to as antigen or as immunogen. Antigen is any agent capable of binding specifically to components of the immune response e. g. B cell receptor (BCR) or B lymphocytes and soluble antibodies. An immunogen is any agent capable of inducing an immune response and is therefore immunogenic.
IMMUNOGENS AND ANTIGENS All immunogens are antigens, but not all antigens are immunogens. A hapten is a compound that by itself, is incapable of inducing an immune response but against which an immune response can be induced by immunization with the hapten conjugated to a carrier. The immune response is directed against various parts of the conjugate including the hapten which can bind antibody molecules.
REQUIREMENTS FOR IMMUNOGENICITY 1 - Foreigness Animals normally do not respond immunologi-cally to self. The more foreign the substance, the more immunogenic it is. In exceptional cases autoimmunity is formed, the immune response is directed against his or her own tissues.
REQUIREMENTS FOR IMMUNOGENICITY 2 - High Molecular weight Small compounds with a M. wt. < 100 Da "e. g. penicillin, aspirin, progesterone" are not immunogenic. Compounds of M. wt. between 1000 -6000 Da "e. g. insulin, ACTH" may or may not be immunogenic. Those of M. wt. > 6000 Da "e. g. albumin" are generally immunogenic.
REQUIREMENTS FOR IMMUNOGENICITY 3 - Chemical Complexity Simple molecules such as homopolymers of amino acids "e. g. a polymer of lysine with a M. wt. of 30, 000 Da" are seldom good immunogen. Copolymers of several amino acids e. g. polyglutamic, alanine and lysine tend to be highly immunogenice. 4 - Degradability Antigens that activate T cells to stimulate immune response must interact with MHC molecules expressed on APCs.
4 - Degradability APCs must first degrade the antigen through antigen processing "enzymatic degradation of antigen" before they can express epitopes on their surface. These epitopes stimulate the activation and clonal expansion of antigen-specific effector T cells. D-amino acids are resistant to enzymatic degradation and are not immunogenic. L- isomers are susceptible to enzymes and are immunogenic. Carbohydrates are not processed or presented, they are unable to activate T cells, but they activate B cells.
REQUIREMENTS FOR IMMUNOGENICITY 5 - Further Requirements for Immunogenicity The genetic makeup "genotype". Genetic control of immune response is controlled by gene mapping within the MHC. The presence of antigen-specific B and T cells.
6 - Dosage and Route of administration Repeated administration of antigen stimulates a strong immune response. Insufficient doses of antigen may not activate enough lymphocytes or because such a dose renders the responding cells unresponsive "tolerance". Subcutaneous administration of antigen elicit the strongest immune responses. Langerhans cells process and present antigen to effector cells. I. V administered antigens are carried to the spleen where they may induce tolerance or If presented by APCs generate and immune response. G. I. T route elicits local antibody responses within the intestinal lamina propria but often produce a systemic state of tolerance. Administration of antigen via intranasal route "respiratory tract" often elicits allergic responses.
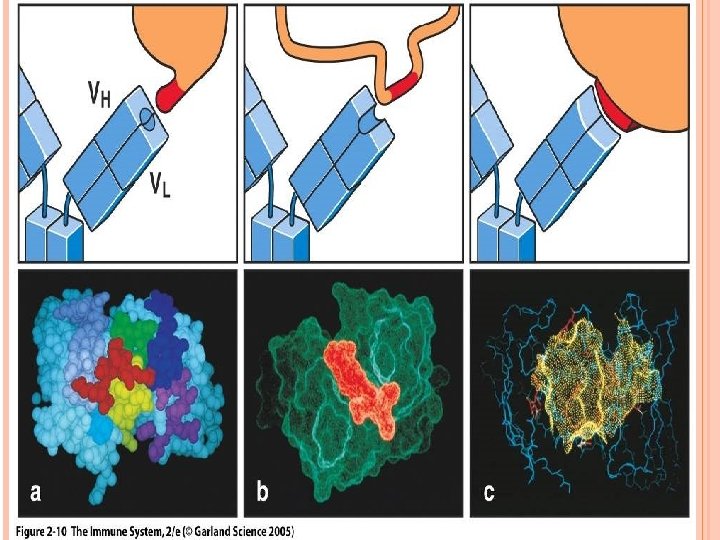
ANTIGENICITY AND ANTIGEN-BINDING SITE An immune response induced by an antigen generates antibodies or lymphocytes that react specifically with the antigen. The portion of the antibody that specifically binds to the epitope is concentrated in several hypervariable regions of the molecule which form the complementarity-determining region "CDR". The size of the epitope that combines with the CDR on a given antibody is approximately equivalent to 5 -7 amino acids. Epitope binds to TCR "8 -12 amino acids" is no covalently associated with MHC proteins of APC. A trimolecular complex is formed "TCR-epitope-MHC"
ANTIGENIC DETERMINANTS (EPITOPES SHAPE)
MAJOR CLASSES OF ANTIGENS 1 - Polysaccharides Immunogenic when associated with protein carries. Glycoproteins are good immunogens. ABO blood groups which are polysaccharides on the surface of the RBCs are examples of antigenicity of polysaccharides. 2 - Lipids are rarely immunogenic. Lipoproteins, glycolipids are immunogenic.
MAJOR CLASSES OF ANTIGENS 3 - Nucleic Acids They are poor immunogens. Nucleoproteins are good immunogens. In SLE anti-DNA antibodies are formed. 4 - Proteins All proteins are immunogenic. Complex proteins are more immunogenic because they contain multiple epitopes.
CROSS REACTIVITY Toxin and Toxoid A toxin that has been modified to loose its toxicity but retains its immunogenecity is called toxoid. Toxoid is used for immunization. Toxoid cross reacts immunologically with toxin. Many epitopes in the toxoid are still reactive. When the antigen used for immunization is different form the one with which the induced immune components are then allowed to react, the terms homologous and heterologous are used.
CROSS REACTIVITY Homologous denotes that the antigen and the immunogen are the same. Heterologous denotes that the substance used to induce the immune response is different from the substance that is then used to react with the products of the induced response.
Cross reacting compounds share antigenic determinants. Some cross-reacting substance are unrelated to each other except that they have one or more epitopes in common. These substances are referred to as heterophile antigens. e. g. human blood group B antigen reacts with antibodies to certain strains of E. coli. The antigens of the microorganisms are referred to as heterophile antigens with respect to the blood group antigen.
Adjuvants: An adjuvant is a substance that when mixed with an immunogen, enhances the immune response against the immunogen. An adjuvant does not confer immunogenecity on haptens. Adjuvant mechanisms include: 1. increasing the biological or immunological half-life of vaccine antigens. 2. increasing the production of local inflammatory cytokines. 3. Improving antigen delivery and antigen processing and presentation by APCs. Adjuvants containing microbial components e. g. mycobacterial extracts are the best adjuvants.