A bioremediciban rsztvev aerob mikroorganizmusok lebontsi tvonalak redox

A bioremediációban résztvevő aerob mikroorganizmusok lebontási útvonalak redox enzimek

Bevezető • A legtöbb szennyező esetén gyors és teljes elbontásukra jellemzően aerob feltételek mellett találunk példát • Az aerob lebontás alapvető feltételei az aerob mikroorganizmusokban: – A mikrobának kapcsolatba kell lépnie a szennyezőanyaggal, pl ha hidrofób vegyületről van szó, először felületaktív anyagot kell termelnie a sejtnek – A támadás egy oxidációs lépéssel kezdődik, az aktiváció és az oxigén beépülése a kulcs reakció, melyet oxigenázok, peroxidázok katalizálnak – Lebontási útvonalak segítségével a vegyület olyan intermedierré alakul, mely beléphet a központi anyagcsere útba pl. a trikarbonsav ciklusba – A keletkező intermedierekből pl. acetyl-Co. A, piruvát, szukcinát biomasszát képez • Számos baktérium, gomba képes a szervesanyagok elbontására (biodegradáció, mineralizáció), átalakítására, vagy úgy, hogy közben szaporodik, vagy kometabolizmussal • Az oxidációs reakciót oxigenázok, peroxidázok, lakkázok katalizálják

Forrás: H-J Rhem, G. Reed: Biotechnology vol 11 b WOLFGANG FRITSCHE and MARTIN HOFRICHTER

Tipikus, a biodegradációban komoly szerepet játszó aerob baktériumok • Ált kemoorganotróf fajok, melyek nagyon sokféle xenobiotikus komponenst képesek szén- és energiaforrásként hasznosítani • Nem várhatjuk, hogy egy baktérium képes legyen minden vegyület bontására, így kevert közösségek segítségével érhetünk el jó eredményt, így alakulhat ki egy erős biodegradatív potenciál • Ez a degradatív potenciál és a környezeti faktorok fogják meghatározni a biodegradáció mértékét • A legdominánsabb és legkönnyebben szelektálható baktériumok környezetünkben a pseudomonasok (tudjuk, hogy a talajban jelenlévő mikroorg-k többségét nem tudjuk elővarázsolni laboratóriumban szaporítással). Szintén jelentősek a Comamonas, Burkholderia, Xanthomonas fajok (G-negatív képviselők). Akár több, mint 100 szerves vegyületet is képesek szénforrásként hasznosítani. • A G-pozitív baktériumok közül a legjelentősebbek a Bacillus és Rhodococcus fajok

Alifások szaporodással kapcsolt bontása • Az alifások és cikloalifások lebontásában a kezdeti lépéshez O 2 -re van szükség • A konkrét lebontási folyamat a szubsztrát tulajdonságaitól, és a lebontásban résztvevő mikroorg enzimkészletétől függ • A hosszabb szénláncú alkánok (C 10 -24) lebontása ált nem gond, a rövidebb láncúak (C<9) viszont toxikusak sok mikroorg számára. • Oxidációjuk terminális vagy diterminális, az első a fő útvonal, alkohol, aldehid és zsírsav keletk, a zsírsavak b-oxidációja során aztán acetil-Co. A képz • Az elágazások ált rontják a biodegr hatékonyságát

n-alkánok oxidációja 1 n-alkán monooxigenáz 2 alkohol dehidrogenáz 3 aldehidrogenáz Forrás: H-J Rhem, G. Reed: Biotechnology vol 11 b WOLFGANG FRITSCHE and MARTIN HOFRICHTER

aromások oxidációja • Aromás vegyületeket minden élő szervezet tud szintetizálni (pl aromás aminosavak, fenolok, kinonok), így nem meglepő, hogy a környezetben szennyezőként megjelenő aromás vegyületeket is képesek bontani. • Az aromások enzimatikusan természetes intermedierekké alakulnak: katekol, protokatekol, melyek a központi anyagcsere útvonalakon alakulnak tovább sejtanyaggá • Az első lépés a benzol gyűrű oxidációja, mely egy hidroxiláció vagy epoxidáció, amit oxigenáz enzimek katalizálnak • Az aromások lebontásáért felelős enzimek többsége katabolikus plazmidokon kódolt, melyek egyik gazdából a másikba át tudnak jutni (önállóan), és széles a gazdaspektrumuk. A legtöbb szennyezet környezetből származó G-negatív talajlakó baktériumból lehet ilyen degradatív plazmidokat kimutatni • Ált az aromások bontásában a lehetséges reakciók száma limitált, hidroxiláció, oxigenolitkus gyűrűhasítás, izomerizáció, hidrolizis jell • Az enzimek indukálható természete, és szubsztrát specifitásuknak köszönhetően a baktériumok nagy degradációs potenciállal rendelk

hidroxiláció Központi intermedierek Forrás: H-J Rhem, G. Reed: Biotechnology vol 11 b WOLFGANG FRITSCHE and MARTIN HOFRICHTER

gyűrűhasítás Forrás: H-J Rhem, G. Reed: Biotechnology vol 11 b WOLFGANG FRITSCHE and MARTIN HOFRICHTER

kometabolizmus • Egy vegyület transzformációja, melyből a sejtnek tápanyag haszna nincs, a reakció a szaporodási szubsztrát jelenlétében működik. Ekkor a vegyület kémiailag módosul • A mikroorg szaporodik a megfelelő szubsztráton, és ‘ok nélkül’ oxidálja a kérdéses vegyületet (koszubsztrát), melyet nem hasznosít, viszont más mikrobák számára tápanyagként szolgálhat • A reakció előfeltétele, hogy a szaporodó sejt termelje a megfelelő enzimet és kofaktorokat (pl. oxigenázok és elektrondonorjaik) • Példák: metanotrófok TCE oxidációja a metán-monooxigenáz enzim segítségével, klór-aromások oxidációja

TCE bontása metanotróf baktériumban: Növekedési szubsztrát CH 4 Cl C CH 4 NADH O 2 H 2 O Metánmonooxigenáz Cl H Cl TCE O Cl NAD+ koszubsztrát C C CH 3 OH Cl Cl C H TCE epoxid Köztes metabolizmus szaporodás távozik a sejtből (nem stabil)

Forrás: R. L. Brigmon: Methanotrophic Bacteria: Use in Bioremediation Westinghouse Savannah River Company, Aiken, SC 29808

+ Fenol szubsztrát 4 Cl-fenol koszubsztrát Forrás: G. A. Hill 1, B. J. Milne 1 and P. A. Nawrocki 1: Cometabolic degradation of 4 -chlorophenol by Alcaligenes eutrophus Applied Microbiology and Biotechnology Volume 46, Number 2 / September, 1996

• Aerob lebontási folyamatokban a központi szerepet az oxigenázok játszák • Alkánok, aromás vegyületek oxidációját katalizálják, melynek eredménye alkoholok (melyek továbbalakulnak aldehidek, epoxidok, karboxilsavak keletk) • Ipari biotechnológiai jelentőség

Oxigenázok 1955 Osamuri Hayaishi - felfedezi az oxigenáz enzimeket dioxigenázok H. S. Mason - felfedezi a fenoláz enzimet 1965 - Irwin Gunsalus - citokróm P 450 bakt. monoox. 1970 David Gibson - aromás szénhidrogén dioxigenázok 1980 - monooxigenázok - lebontó útvonalak tanulmányozása sok kutatócsoport - lebontási útvonalak feltérképezése Ananda Chakrabarty - katabolikus plazmid transzfer Pseudomonas törzsek között
 •")
Oxigenázok • Aerob mikroorganizmusokban gyakori, de eukarióta szervezetekben is (pl Citokróm P 450) • Működésükhöz molekuláris oxigénre és kofaktorokra szükség van • Szerkezetükben, működésükben, kofaktor igényükben a különböző oxigenázok eltérhetnek egymástól • Általában több alegységesek, fémet is tartalmaznak, melynek szerepe a dioxigén aktiválásában van. Főleg vasat, de van mangán, réz, kobalt tart-ú is • A katalizált reakció energianyerő, a felszabaduló erg a szubsztrát szerkezetének destabilizálására fordítják • Iparilag hasznosíthatóak, genetikailag módosított enzimekkel jobb hatékonyság, szélesebb szubsztrátspektrum hozható létre

Az aromás gyűrű szerkezet lebontási útvonalában a Prokarióta és Eukarióta oxigenázok katalizálta reakciók első lépcsője

Oxigenázok • Eredetüket tekintve sokszor nagyon eltérő szervezetekből származnak, mégis lehetnek nagyon hasonlóak felépítésüket, szekvenciájukat, specifitásukat tekintve, vagy nagyon eltérőek is lehetnek • Hatékonyságuk összefüggésben szubsztrátspecifitásukkal • Gyakran plazmidon kódoltak • Két nagy csoportra oszthatók: – Monooxigenázok – Dioxigenázok

I. Monooxigenázok • Azokat az oxigenázokat, melyek a dioxigén egyik atomjának a szubsztrátba beépülését katalizálják monooxigenázoknak nev. Az oxigén molekula másik atomja redukálódik és vízmolekulává alakul, melyhez elektrondonorként NAD(P)H-t igényel, vagy magát a szubsztrátot használja • Többségük egykomponensű enzim, de előfordul multikomponensű is pl. fenol-, toluol monooxigenáz Csoportosításuk: 1. aromás gyűrű monooxigenázok (fenol 2 -hidroxilázok, flavoprotein monooxigenázok) 2. alkil csoport hidroxilázok (alkán hidroxilázok, metán monooxigenáz) 3. citokróm P-450 család – Multikomponensű enzimkomplex: egy terminális hidroxiláz és egy v több elektrontranszport komponens (főleg emlősökben) 4. aromás aminosav hidroxilázok (Eukariótákban pl triptofán hidroxiláz)
ba építik. Csoportosításuk:")
II. Dioxigenázok • A dioxigenázok az O 2 mindkét atomját a szubsztrát(ok)ba építik. Csoportosításuk: 1. Az aromás gyűrűre oxigént építő - mindig orto pozícióban dihidroxilálnak, katekol származék keletkezik - funkcionálisan két rész: egy hidroxiláz komponens és egy v több e- transzport komponens - A reakcióhoz NAD(P)H-ra is szükség van. 2. Az aromás gyűrű hasításában résztvevő dioxigenázok - A gyűrűt hasító dioxigenázok a gyűrű felnyitását katalizálják, melyhez külső reduktánsra nincs szükség - 2 v. több –OH csoportot tartalmazó aromás gyűrűk a szubsztrátjaik

II. Dioxigenázok 2. Az aromás gyűrű hasításában résztvevő dioxigenázok - Ha a 2 –OH csop. egymáshoz visz. helyzete orto, akkor több pozícióban is hasíthatják a gyűrűt: megkülönböztetünk 2, 3 -; 3, 4 -; 4, 5 -gyűrű hasító dioxigenázokat: a, 2, 3 -; 4, 5 -dioxigenázok a katekolokat extradiol/meta, b, 3, 4 -dioxigenázok intradiol/orto helyzetben hasítják - Ha a 2 –OH csop. egymáshoz visz. helyzete para pl. gentizát esetén a karboxil csoport és a mellette lévő hidroxil csoport között hasít

II. Dioxigenázok NADH+H+ O 2 NAD+ YH

A hasító helytől függően eltérő intermedierek keletkeznek a további átalakítás során

Gombák degradatív képessége • A gombák széles körben elterjedtek környezetünkben (talajban, édes-, tengervízben, hulladékban, avarban, állati-, növényi maradványokon, trágyában, élő szervezetekben) • Szaprofita, lebontó, szimbionta, parazita • Fontos degradatív kapacitással és képességekkel rendelkeznek, többek között a veszélyes anyagok elbontásában is szerepet játszanak • mikroméretű gombák – élesztők, penészek - és Basidiomyceták élesztő penész Phanerochaete sp

Mikroméretű gombák jelentősége a környezetvédelemben • E csoportot az élesztők és penészek alkotják • Képesek különféle szervesanyagot lebontani, szén és energia forrásként hasznosítani pl alifás szénhidrogének, aromások • Legjobban a C 10 -20 szénhidrogéneket kedvelik, de oxidálják a hosszabb szénláncokat is, viszont a rövid láncúak toxikusak számukra. Tipikus alkánbontók: – Élesztők közül példák: Candida lipolytica, Rhodotorula rubra, … – Penészek közül: Cunninghamella blakesleeana, Aspergillus niger, Penicillium frequentans, … • Mivel hidrofób vegyületek bontásáról van szó, e gombák is képesek felületaktív anyagot termelni • Gombákban a lebontási folyamat főleg monooxigenáz enzimkomplex katalizálta terminális oxidációval indul, mely során alkohol keletk a további bontást peroxiszomális enzimek (oxidázok, katalázok) végzik (b-oxidáció). • Képesek hasznosítani az alkéneket is, de az elágazó láncú alkánokat, cikloalkánokat nem szeretik

Mikroméretű gombák jelentősége a környezetvédelemben • Sok gomba képes hasznosítani aromás vegyületeket is szén és energiaforrásként, de főleg kometabolizmussal alakítják át • Az aromás gyűrűt orto-hasító útvonalon bontják, amihez szükséges két OH csoport orto helyzetű jelenléte, amit hidroxiláz enzimek segítségével kapcsol a gyűrűre. Ezek az enzimek. NAPH függő monooxigenázok orto és para helyzetben tudnak hidroxilálni. A felhasadt vegyület továbbalakul és belép a trikarbonsav ciklusba, így biomassza, CO 2 és víz keletkezik. Az elbontandó vegyület felvételét specifikus energia-függő felvételi rendszerrel biztosítja. • A gyűrűhidroxiláló és gyűrűhasító enzimeik előnye, hogy relatíve nem specifikusak, így többféle vegyületet is támadnak pl. halo-, nitroaromások… • Kometabolikus reakcióban számos vegyületet képesek átalakítani, pl PAH-ok, bifenilek, dibenzofuránok… • Degradatív képessük miatt jelentősek a szén-ciklusban, és a talaj öntisztító képességében

Basidiomyceta gombák jelentősége a környezetvédelemben • Makroszkópikus méretűek, főleg az avarban, a talaj felső rétegében, erdőkben, legelőkön és lignocellulóz tartalmú anyagokon találjuk meg őket • A cellulózt védő lignin elbontásában óriási szerepük van Phanerochaete chrysosporium

Ligninolitikus enzimrendszer • A lignin a cellulóz mellett a növények anyagainak egyik fő képviselője, a növény támasztórendszerét erősíti, a rigiditásért felelős • Gátat jelent a mikrobákkal szemben, megvédi a bontható poliszaharidokat (cellulóz, hemicellulózok) • Heterogén polimer, fenilpropán egységekből épül fel, melyek sokféle kovalens kötéssel kapcsolódnak egymáshoz • A kötések típusa és heterogenitása miatt hidrolitikusan nem bontható (ami egyébként a polimerek bontására jellemző) • A kötés sokféleség miatt igen meglepő, hogy az evolúció során egyes fajok, basidiomyceta gombák képessé váltak e polimer bontására • a lignint szelektíven bontják, a cellulózt (fehér rostokként látszanak) nem bántják, ezért fehér rothasztó gombáknak is nevezzük őket: – Fákat támadók: Trametes versicolor, Phanerochaete chrysosporium, Pleurotus ostreatus – Avar rothasztók: Stropharia rugosoannulata, Mycena galopus, Agrocibe praecox

Ligninolitikus enzimrendszer • Noha a lignin bontható, nem elsődleges szénforrása a gombáknak, szaporodásukhoz a jelenlévő hemicellulózokat hasznosítják • A lignin bontásban több enzim kooperál: mangán peroxidáz (Mn. P), lignin peroxidáz (Li. P), lakkáz – nem specifikus reakciók. A peroxidázok működéséhez peroxidra szükség van, a lakkáz O 2 –t igényel • A lakkáz és a Li. P az aromás szubsztrátot támadja közvetlen oxidációval szabad gyököt képez, míg a Mn. P közvetve hat egy kelált Mn 3+ ionon keresztül, mint alacsony molekulatömegű redox mediátor (a funkciója Mn 2+-ből, ami a szubsztrátja az enzimnek Mn 3+-at képez, mely nagyon reaktív). A Mn. P-nak szüksége van Mn-keláló ágensre pl. oxalát, malát. A kelált Mn elég kicsi molekula komplex ahhoz, hogy bejusson a lignocellulóz hálóba, és ott reagál a fenolokkal • A kezdeti enzimtámadás eredménye vízoldékony lignin fragmentek, melyek továbbalakulnak a lignolitikus enzimek segítségével
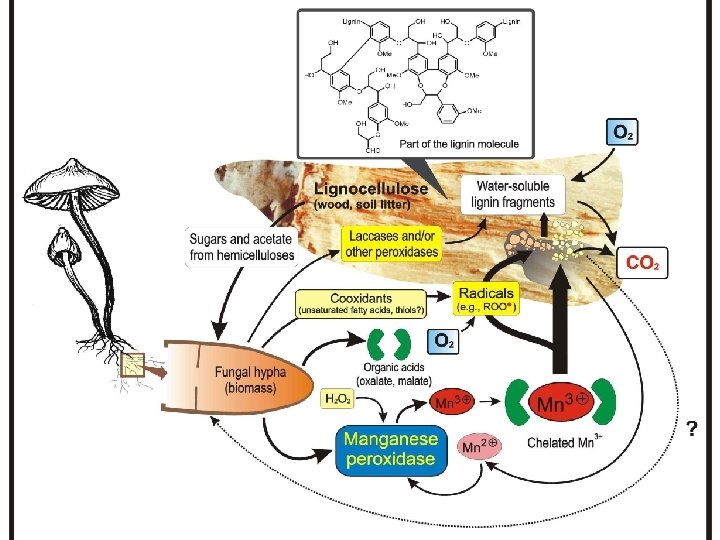

Manganese Peroxidase Three basic monomers of lignin

Ligninolitikus enzimrendszer • A Basidiomyceta gombák lignolitikus enzimrendszere az erős reaktív szabadgyökökkel tört depolimerizációs mechanizmusuk alapján alkalmasak szerves szennyezőanyagok biodegradációjára • Az extracelluláris, nem specifikus, nem-sztereoszelektív tulajdonságuk miatt számos toxikus, bontásnak ellenálló vegyület támadására képesek • Amiért mégsem ezek a legjelentősebb rendszerek a természetben a biodegr során, azért van, mert kompeticiós képességük kicsi. A korhadást okozó gombák a talajban csak rövid ideig képesek túlélni (csak a fás körny-t szeretik) • Ennek ellenére bizonyos esetekben csak ezek vethetők be megfelelő körülményeket biztosítva • Összességében nem mondhatjuk, hogy a baktériumok/ vagy éppen a gombák a jobb lebontók, az adott körülmény határozza meg melyiket használjuk

Anaerob kooperáció • A szervesanyagok, főleg aromások esetén a funkciós csoportok (pl. halo-, nitro-, szulfo-) számának növekedésével egyre hosszabb ideig lesznek jelen a körny-ben • Minél inkább elektrofil egy molekula, annál kevésbé támadható elektrofil támadással pl. oxigenázokkal (- aerob mikroorg) • A természetben is pl halogén tart-ú vegyületek esetén főleg anaerob mikroorg reduktív támadása jellemző, általános a természetben, legtöbb esetben konzorciumok végzik, de ritka a teljes elbontása az adott vegyületnek, A keletkező termék az aerobok számára is legtöbb esetben már elfogadható, így mineralizálódik, vagy továbbalakul ezek által
- Slides: 33