Wielkoskalowa charakterystyka genomw Chromatyna Chromatyna wypenia jdro komrkowe







 Olins & Olins, 1973 http: //users. rcn. com/jkimball.")


")


















 nagromadzają się w życiu osobniczym • Significant")
 Lcyc kontroluje symetrię grzbietowobrzuszną kwiatu;")







 • HDAC –")









 chromatyny Główne typy kompleksów remodelujących chromatynę SWI/SNF Snf 2 Swp 73")

 chromatyny Jeden z mechanizmów remodelingu: przesunięcie oktameru histonów wzdłuż nici DNA")
 chromatyny Pozycja skrajna Pozycja centralna")


 i zamkniętych (B) promotorów. • Typowa mapa")

 poprzez mitozę, nie wymagające zmian w sekwencji")









 kwitna wcześnie Jesień Zima Wiosna FLC jest")




- Slides: 74
Wielkoskalowa charakterystyka genomów Chromatyna
Chromatyna wypełnia jądro komórkowe
23 ludzkie chromosomy w jądrach fibroblastów we wczesnej profazie
Maksymalny stopień kondensacji DNA osiąga w chromosomach
Białka histonowe
Chromatyna trawiona nukleaza, oryginał z pracy Hewish and Burgoyne 1973 Dean Hewish, 1973 Leigh Burgoyne, 1973
Trawienie chromatyny MNazą - drabinka nukleosomowa
„Sznur korali” (‘beads on the string’) Olins & Olins, 1973 http: //users. rcn. com/jkimball. ma. ultranet/Biology. Pages/N/Nucleus. html
Roger Kornberg w 1974 r. zaproponował model, w którym DNA owinięty jest wokół rdzenia histonowego tworząc nukleosom NUKLEOSOM JEST PODSTAWOWĄ JEDNOSTKĄ STRUKTURALNĄ CHROMATYNY
Fałd histonowy
Złożenie fałdów (hand shake)
Konserwowane elementy na obrzeżu oktameru
Oktamer – oddziaływanie z DNA
Mutacje SIN znoszą oddziaływania oktameru z DNA
Składanie nukleosomu
Zaginanie i zwijanie DNA na oktamerze • Ściśle zwinięta lewoskrętna superhelisa ma ok. 80 pz na zwój i skok superhelikalny ok. 27, 5 A.
Ekspozycja miejsc w helisie DNA na oktamerze
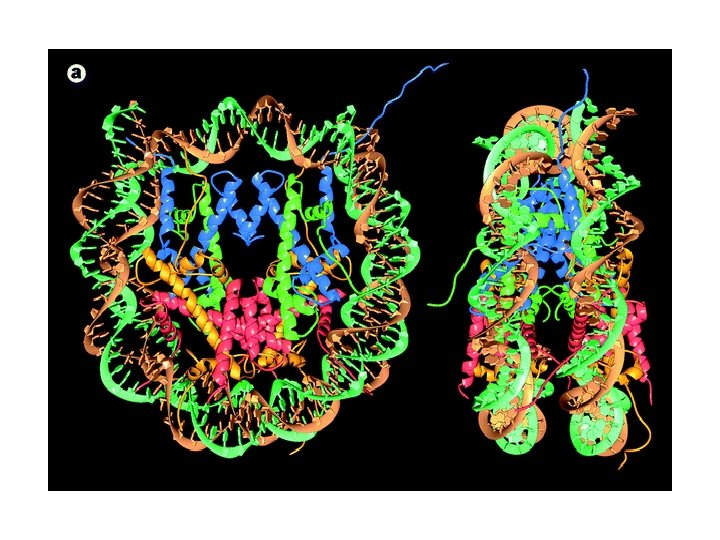
Struktura krystalogtaficzna cząstki rdzeniowej nukleosomu Karolin Luger Timothy Richmond Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Nature 1997 Sep 18; 389(6648): 251 -60
Schemat nukleosomu
Nukleosomy - terminologia 8 histonów: Po dwa każdego z: H 2 A H 2 B H 3 H 4 Gräff and Mansuy (2008). Behav Brain Res.
Organizacja chromatyny Podwójna helisa DNA 2 nm Histony + DNA. “Koraliki na sznurku” f Komórka roślinna 11 nm Włókna nukleosomowe 30 -nm (solenoid) 30 nm Włókna połączone z macierzą jądrową 300 nm Nuclear Matrix
Umiejscowienie H 1 w nukleosomie
Regulacyjna rola chromatyny
Struktura a funkcja chromatyny
Zmiany struktury chromatyny • modyfikacje DNA • modyfikacje potranslacyjne histonów • wyspecjalizowane warianty histonów • ATP-zależna przebudowa (remodeling) chromatyny
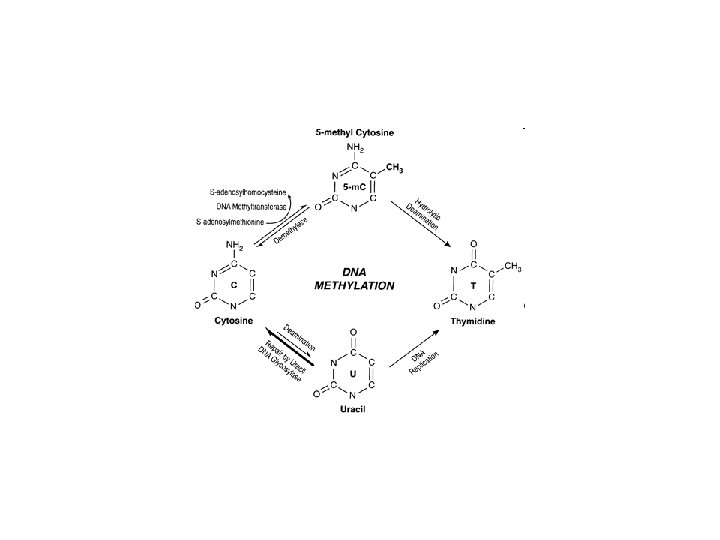
Metylacja DNA, Metylomy
Metylacja DNA zamienia cytozynę w 5 -metylo cytozynę
Metylacja DNA ma znaczenie biologiczne Arabidopsis 46 -dni od wysiania e as r sfe n a r NH 2 N O ylt h t e NH 2 M CH 3 N N ~ cytosine O N ~ 5 -methylcytosine WT met 1/cmt 3 Xiao, et al. (2006). The Plant Cell. 18: 805 -814
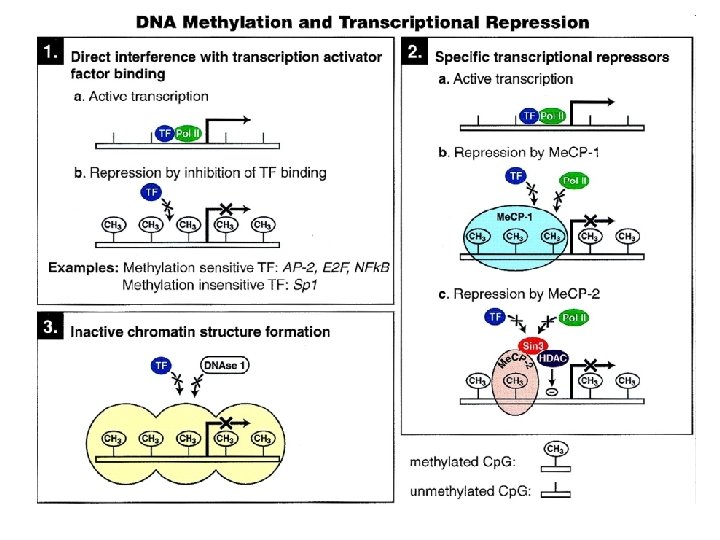
Metylacja DNA u eukariontów • Nie jest uniwersalna, występuje u ssaków i roślin kwiatowych • Jest zmienna gatunkowo, tkankowo i chromosomowo • Rozpoznawana przez rodzinę białek zawierających domenę (MBD) wiążącą metylowany DNA • Ściąga kompleksy białkowe indukujące zmiany w lokalnej strukturze chromosomów • Wyłącza ekspresję genów • Zakłócenia w normalnym wzorze metylacji DNA są niemal powszechnie wykrywane w nowotworach
U bliźniąt monozygotycfznych różnice epigenetyczne (metylacja DNA) nagromadzają się w życiu osobniczym • Significant DNA methylation changes are indicated as thick red and green blocks in the ideograms. The 50 -year-old twin pair shows abundant changes in the pattern of DNA methylation (green=hypermethylation and red=hypomethylation), 3 -year-old twins have a very similar DNA methylation (yellow). • ". . . We found that, although twins are epigenetically indistinguishable during the early years of life, older monozygous twins exhibited remarkable differences in their overall content and genomic distribution of 5 -methylcytosine DNA and histone acetylation, affecting their gene-expression portrait. These findings indicate how an appreciation of epigenetics is missing from our understanding of how different phenotypes can be originated from the same genotype. " • Proc Natl Acad Sci U S A. 2005 Jul
Przykład ewolucyjnego efektu epimutacji (zmiany we wzorze metylacji DNA) Lcyc kontroluje symetrię grzbietowobrzuszną kwiatu; u mutanta nieaktywny z powodu silnej, dziedziczonej metylacji From Cubas et al 1999, Nature 401: 157 -161
Epigenetyczna kontrola czasu kwitnienia Przedłuzony okrers chłodu Rozwój generatywny Rozwój wegetatywny Jesień Zima Wiosna Niektóre rośliny wymagają przedłużonego okresu chłodu (wernalizacji) – w trakcie zimy, zanim będą mogły zakwitnąć.
Transpozony • Fragments of DNA that can insert into new chromosomal locations • Some copy themselves and increase in number within the genome • Responsible for large scale chromosomal rearrangements as well as single-gene mutagenic events
U roślin program epigenetyczny wycisza transpozony i chroni integralość centromerów
Transpozony mogą powodować wyłączenie lub niestabilność alleli Gen biosyntezy barwnika Allel dziki Ziarniak barwny Gen rozbity przez transpozon Allel zmutowany Ziarniak bezbarwny Wycięcie transpozonu powoduje niestabilność allelu Allel niestabilny Ziarniak częściowo zabarwiony
Metylacja DNA jest niezbędna do wyciszania transpozonów BLUE = Gene density RED = Repetitive element density Utrata funkcji met 1 lub ddm 1 (decrease in DNA methylation 1) mutanty maja niedometylowany DNA GREEN = Methylated DNA BROWN = Methylated DNA in a met 1 mutant Zhang, et al. (2006). Cell 126: 1189 -1201
Aktywacja transpozonów w mutancie ddm 1 indukuje mutacje Dziki Po 1 pokoleniu Po 3 pokoleniach Po 5 pokoleniach Po wyłączeniu DDM 1, fenotyp rośliny staje się coraz bardziej nienormalny, w miarę akumulowania indukowanych przez transpozony. Kakutani, et al. (1996). PNAS. 93: 12406 -12411
Lokalizacja ogonów histonowych w nukleosomie H 4 H 3 H 2 A H 2 B
Enzymy modyfikujące histony i DNA • HAT – acetylotransferazy histonów (bromodomena) • HDAC – deacteylazy histonów • HMT – metylotransferazy histonów (chromodomena). CMT 3 – DNA metylotransferaza (chromodomena)
Przykład kowalencyjnej modyfikacji aminokwasów acetylacja lizyny
Modyfikacje potranslacyjne białek histonowych Kozaurides 2007
Elektroforetyczny dowód acetylacji histonów
Efekt acetylacji ogonów histonowych
Mechanizm działania modyfikacji potranslacyjnych białek histonowych Ø Działanie bezpośrednie: zmiany w oddziaływaniach histon-DNA i histon-histon Ø Działanie pośrednie: rekrutacja białek rozpoznających określone modyfikacje Kozaurides 2007
Fenotyp mutantów w metylotransferazie histonów Mutanty są pozbawione metylacji w H 3 K 36. Dong, et al. (2008). Biochemical and Biophysical Research Communications. 373: 659 -664
Modyfikacje a struktura chromatyny H 3 K 9 di. ME euchromatyna heterochromatyna
Warianty histonów – H 2 A
Warianty histonów - rozmieszczenie w chromosomach
ATP-zależna przebudowa (remodeling) chromatyny Główne typy kompleksów remodelujących chromatynę SWI/SNF Snf 2 Swp 73 Snf 5 Swi 3 ISWI Mi 2
Działanie kompleksów remodelujących chromatynę ATP DDM 1 Kingston & Narlikar, 1999 Brzeski & Jerzmanowski, 2003
ATP-zależna przebudowa (remodeling) chromatyny Jeden z mechanizmów remodelingu: przesunięcie oktameru histonów wzdłuż nici DNA
Przykład „przebudowy” (remodelingu) chromatyny Pozycja skrajna Pozycja centralna
Pozycjonowanie nukleosomów
Nukleosomy zajmują określone miejsca w chromatynie • • • A. Schemat ułożenia nukleosomów w chromatynie, pojęcie długości powtórzeń nuklesomowych. B. Silnie zlokalizowane i rozmyte nukleosomy. • C. Nukleosomy a miejsca wiązania czynników transkrypcyjnych. • G. Arya et al. jbsd 2010 •
Nukleosomy a transkrypcja • Schemat otwartych (A) i zamkniętych (B) promotorów. • Typowa mapa pozycyjna nukleosomów na otwartym promotorze pokazuje wyraźne wyróżnienie locus genowego przez obecność rejonów 5’ i 3’ wolnych od nukleosomów (NDR). • (TSS) - miejsce startu transkrypcji. • 5’ NDR sąsiaduje z silnie związanymi nukleosomami. • G. Arya et al. jbsd 2010
Dziedziczenie epigenetyczne • Dziedziczne modyfikacje funkcji genów nie związane ze zmianami w sekwencji DNA • Mechanizmy epigenetyczne są związane z modulacją chromatyny i obejmują: modyfikacje histonów, metylację DNA, przebudowę nukleosomów (remodeling), system RNAi.
Epigenetyka Dziedziczenie właściwości (np. wzoru ekspresji genów) poprzez mitozę, nie wymagające zmian w sekwencji DNA. Genetyka Epigenetyka Allis, et al. (2007). Epigenetics, overview and concepts. CSHL.
Dlaczego sklonowany kopt nie jest dokładnie taki, jak oryginał? Rainbow Klon Rainbow
Modyfikacje histonów Biotinylation ADP-ribosylation k 6 k 11 * * * Gräff and Mansuy (2008). Behav Brain Res.
Po-translacyjne modyfikacje histonów w nukleosomie wg. B. Turner, Cell 2002
Przykład przeciwstawnych funkcji modyfikacji histonów • H 3/H 4 Ac. Lys/H 3 met. Lys 4(H 3 K 4) związane z rejonami aktywnej transkrypcji H 3/H 4 Lys+/H 3 met. Lys 9 (H 3 K 9) związane z rejonami wygaszonej transkrypcji.
Euchromatyna i heterochromatyna Euchromatyna Heterochromatyna • Mniej skondensowana • Silnie skondensowana • Głównie w ramionach chromosomów • Głównie w centromerach i telomerach • Zawiera unikatowe sekwencje • Zawiera powtórzone sekwencje • Bogata w geny • Uboga w geny http: //www. nyas. org/
Determinanty aktywnej i wyciszonej chromatyny
Znaczniki epigenetyczne związane ze stanami chromatyny ATP-zależna przebudowa chromatyny ATP-dependent chromatin remodeling
X chromosome inactivation involves epigenetic silencing In female mammals, one copy of the X chromosome in each cell is epigenetically inactivated. Fur color in cats is determined in part by orange, an X-linked gene. A female cat that is heterozygous for the orange gene shows black and orange patches, corresponding to which X chromosome is active.
Program epigenetyczny wspomaga przejścia rozwojowe u roślin Przejście z fazy embrionalnej do wegetatywnej Rozwój embrionalny Przejście z fazy wegetatywnej w generatywną Rozwój wegetatywny Rozwój generatywny
Mutanty w genie FLOWERING LOCUS C (FLC) kwitna wcześnie Jesień Zima Wiosna FLC jest inhibitorem kwitnienia; brak FLC znosi konieczność wernalizacji.
FLC hamuje FT, aktywator kwitnienia Roślina dzika FL C Gen FT Związanie FLC hamuje transkrypcję FT Roślina z mutacją flc Gen FT F T
FLC jest wyciszany w trakcie wernalizacji Po 40 dniach w 4°C , FLC nie jest wyrażany. 10 dni po powrocie do 22°C ekspresja FLC jest wciąż wyłączona. Jesień FLC transkrybowany Zima Wiosna FLC wyciszony Sung, et al. (2004). Nature. 427: 159 -164.
FLC jest regulowany przez modyfikacje epigenegtyczne H 3 k 4 me H 3 K 36 me H 3 K 9 ac H 314 ac H 3 k 9 me 2 H 3 k 27 me 2 Zimno Jesień FLC transkrybowany Zima Wiosna FLC wyciszony
Kompleksy VIN 3 i PRC 2 epigenetycznie wyłączają FLC PRC 2= Polycomb Represive Complex 2) VIN 3 PRC 2 (łącznie z VIN 3) Jesień FLC transkrybowany Zima Wiosna FLC wyciszony