CALCOLO DELLE FREQUENZE GENICHE PER GENI LEGATI AL

/2 N")



 AAx. AA=Sp 4=Sp 2")




 AA = (1 -F) p 2 Aa = (1 -F)2")
 p 2 =")




6 = 4(1/64) = 1/16 1/2")
8 = 4(1/256) = 1/64")
5 = 4(1/32) = 1/8")

- Slides: 22
CALCOLO DELLE FREQUENZE GENICHE PER GENI LEGATI AL SESSO GLUCOSIO-6 -FOSFATO G 6 PD 6 -FOSFO-LATTONE A (110%) ALLELE NORMALE B (100%) A- (Africa) ALLELE NORMALE ALLELI B- (Mediterraneo) Gd. Med Gd. BGd. A- Gd. Med
FEMMINE XGd. BXGd. Med MASCHI XGd. BY FEMMINE Alleli codominanti ± p(1 -p)/2 N XGd. Med. Y MASCHI Conteggio diretto ± p(1 -p)/N
POPOLAZIONE AFRICANA (L’allele non deficitario può avere delle varianti di mobilità elettroforetica- più veloci/più lente) B 67 • • A 13 A 8 Gd. B = 67/90 =0. 744± 0. 0021 Gd. A = 13/90 =0. 144± 0. 0370 Gd. A- = 8/90 = 0. 090± 0. 0302 Gd 93% = 2/90 =0. 022± 0. 0155 93% 2 N 90
OROMO Maschi A 6 B 30 36 Femmine AB 5 B 40 45 AMHARA OROMO 4 42 46 Gd. A=0. 1667± 0. 0621 Gd. B=0. 8333 AMHARA Gd. A=0. 0870± 0. 0416 Gd. B=0. 9130 OROMO 3 37 40 Gd. A=0. 0556± 0. 0242 Gd. B=0. 9444 AMHARA Gd. A=0. 0375± 0. 0212 Gd. B=0. 9625
ACCOPPIAMENTO ASSORTATIVO • Accoppiamento assortativo positivo completo incompleto • Accoppiamento assortativo negativo completo incompleto
Accoppiamento assortativo positivo incompleto Frazione S della popolazione (accoppiamento assortativo) AAx. AA=Sp 4=Sp 2 Aax. Aa=S 4 p 2 q 2=S 2 pq aaxaa=Sq 4=Sq 2 Frazione 1 -S della popolazione (accoppiamento casuale) AAx. AA=(1 -S)p 4 AAx. Aa=(1 -S)4 p 3 q AAxaa=(1 -S)2 p 2 q 2 Aax. Aa=(1 -S)4 p 2 q 2 Aaxaa=(1 -S)4 pq 3 aaxaa=(1 -S)q 4
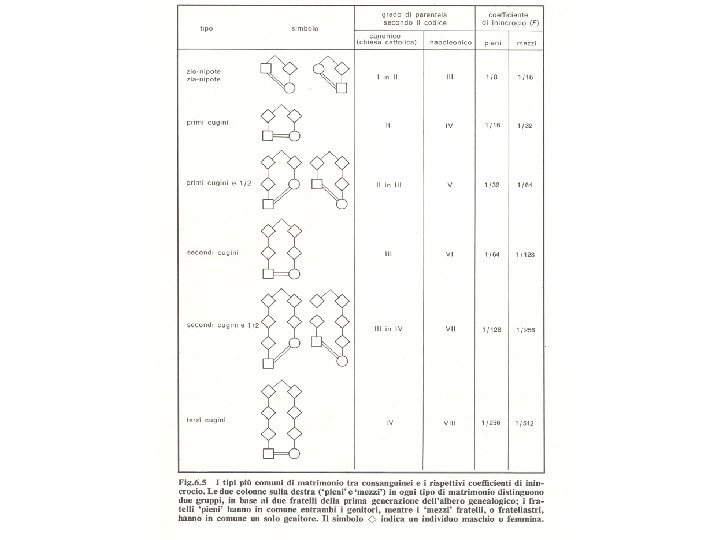
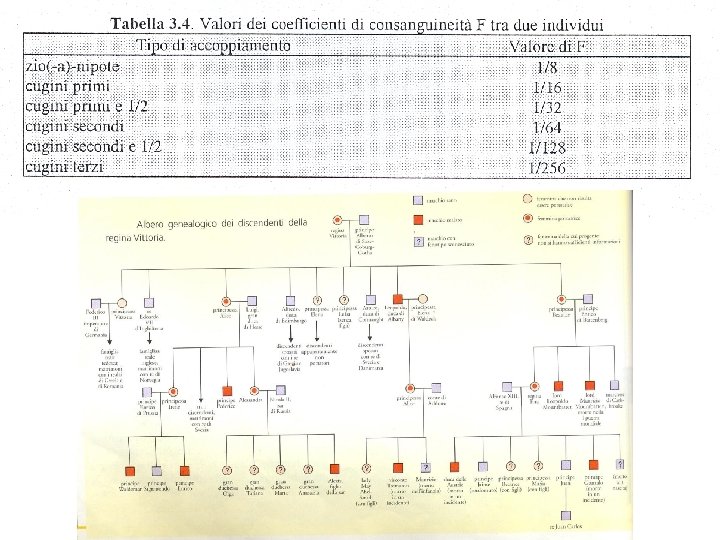
Accoppiamento consanguineo Incrocio tra parenti prossimi inincrocio Molto usato in agricoltura e nell’allevamento degli animali per ottenere individui con particolari caratteristiche Nell’uomo motivi sociali e culturali riducono, ma non eliminano del tutto, l’incidenza del matrimonio tra consanguinei I matrimoni consentiti sono quelli tra primi cugini, tra secondi cugini e tra zio/a e nipote
La principale conseguenza genetica aumento della frequenza degli omozigoti possibilità che si manifestino geni recessivi rari Geni recessivi rari spesso dannosi aumento del carico genetico nella popolazione (parte della variabilità genetica che in accoppiamento casuale rimane nascosta) Comunità geograficamente o socialmente isolate alte frequenze di matrimoni tra parenti ”isolati genetici” “rottura degli isolati”
Accoppiamento consanguineo tipo particolare di unione assortativa 2 individui che hanno almeno 1 antenato comune alleli uguali per discesa FXY = probabilità che 1 individuo riceva, per un certo locus, dai genitori 2 alleli uguali per discesa Somiglianza genetica tra due individui (X e Y) è misurata dal coefficiente di consanguineità Alleli uguali in stato alleli uguali che non derivano da un antenato comune, o meglio da un antenato comune identificabile perché molto lontano nel tempo Il coefficiente di consanguineità tra 2 individui è anche la probabilità che ha l’individuo nato tra i 2 di essere omozigote a 1 locus per 2 alleli uguali per discesa Coefficiente di consanguineità = coefficiente d’inincrocio = coefficiente di consanguineità tra i suoi genitori FXY = FI
Locus con 2 alleli codominanti A e a (frequenza p e q tali che p + q = 1) In accoppiamento casuale AA = p 2 (omozigote alleli uguali solo in stato) Nel caso di accoppiamento consanguineo la popolazione si suddivide in 2 sottopopolazioni: una che pratica accoppiamento consanguineo (proporzione pari a F) una che pratica l’accoppiamento casuale (proporzione pari a 1 F) Nella prima frazione A (p)x. A (p) = 1 p 2 = 1 =1 p=1 AA = Fp aa = Fq La somma delle frequenze dei 2 omozigoti Fp +Fq = F(p+q) = F
Nell’altra frazione (accoppiamenti casuali) AA = (1 -F) p 2 Aa = (1 -F)2 pq aa = (1 -F) q 2 In questo caso la somma delle frequenze (1 -F) p 2 + (1 -F)2 pq + (1 -F) q 2 = (1 -F)( p 2 + 2 pq + q 2) = (1 -F)(p + q)2 = 1 -F
Nell’intera popolazione le frequenze saranno AA = Fp + (1 -F) p 2 = = Fp + p – Fp – pq +Fpq = = p – p(1 -p) + Fpq = = p –p + p 2 + Fpq = = p 2 + Fpq omozigoti Aa = 2 pq(1 -F) eterozigoti↓ aa = p 2 + Fpq
La consanguineità non altera le frequenze alleliche. Nella generazione successiva la frequenza di A p’ = p 2 + Fpq + (1 -F)pq = = p 2 + Fpq + pq-Fpq = p 2 + pq = p(p + q) = p analogamente q’ = q Se F rimane costante di generazione in generazione la popolazione si mantiene in equilibrio → equilibrio di Wright
Calcolo coefficiente di consanguineità con frequenze alleliche La consanguineità determina una riduzione degli eterozigoti La stima del livello viene fatta confrontando la popolazione dei genotipi eterozigoti osservati con quella attesa nell’ipotesi di accoppiamento casuale Popolazione con un certo grado di consanguineità Locus a 2 alleli A (p) e a (q) p+q=1 H = frequenza eterozigoti osservati H 0 = frequenza degli eterozigoti nella popolazione in accoppiamento casuale = 2 pq (Hardy-Weinberg) F= F misura la riduzione di eterozigosità che ha una popolazione che pratica un certo livello di accoppiamento consanguineo rispetto a una popolazione con le stesse frequenze alleliche e che pratica l’accoppiamento casuale
Calcolo coefficiente di consanguineità con frequenze alleliche La consanguineità determina una riduzione degli eterozigoti La stima del livello viene fatta confrontando la popolazione dei genotipi eterozigoti osservati con quella attesa nell’ipotesi di accoppiamento casuale Popolazione con un certo grado di consanguineità Locus a 2 alleli A (p) e a (q) p+q=1 H = frequenza eterozigoti osservati H 0 = frequenza degli eterozigoti nella popolazione in accoppiamento casuale = 2 pq (Hardy-Weinberg) F= F misura la riduzione di eterozigosità che ha una popolazione che pratica un certo livello di accoppiamento consanguineo rispetto a una popolazione con le stesse frequenze alleliche e che pratica l’accoppiamento casuale
ab Probabilità che trasmetta allele a ai figli Generazione filiale: 1/4 ac 1/4 ad 1/4 bc 1/4 bd cd 1/2 Y X FI=FXY 1/2 Probabilità che figlio trasmetta lo stesso allele a ai figli Z Probabilità che la madre trasmetta l’allele a all’individuo Z è (1/2)4 Considerando tutti e 4 gli alleli FI = 4 (1/2)4 = 4(1/16) = 1/4
PRIMI CUGINI 1/2 1/2 FI = 4(1/2)6 = 4(1/64) = 1/16 1/2
SECONDI CUGINI 1/2 1/2 FI = 4(1/2)8 = 4(1/256) = 1/64
ZIO/A E NIPOTE 1/2 1/2 1/2 FI = 4(1/2)5 = 4(1/32) = 1/8
FRATELLI GERMANI ab cd ef 1/2 ac bc ce cf ad bd de df 1/2 FI = 2(1/2)4 = 2(1/16) = 1/8