Vezani geni genetike rekombinacije i mapiranje hromozoma mejoza

Vezani geni, genetičke rekombinacije i mapiranje hromozoma

mejoza interhromozomske rekombinacije • nezavisnim rasporedjivanjem hromozoma u gamete, uvek je rekombinantna klasa zastupljena sa 50% verovatnoće, odnosno, • nezavisno rasporedjivanje daje jednaku verovatnoću parentalnih i neparentalnih gameta u potomstvu • važi za gene na različitim hromozomima, ali i za lokuse jako udaljene na istom hromozomu

grupe vezanih gena - segregiraju zajedno u gamete. aleli odredjene grupe imaju šansu da ostanu zajedno-vezani su.

Mejotičke rekombinacije - mogu da stvaraju haploidne genotipove potomaka koji se razlikuju od haploidnih parentalnih genotipova. intrahromozomalne

krosing-over u mejozi. Potomstvo dobija po jedan homologi hromozom od svakog roditelja Razmena između delova homologih parentalnih hromozoma daje gamete sa različitom kombinacijom alela na hromozomima od roditeljskih

Rekombinacije između gena na homologim hromozomima nastaju krosingoverom nesestrinskih hromatida u mejozi
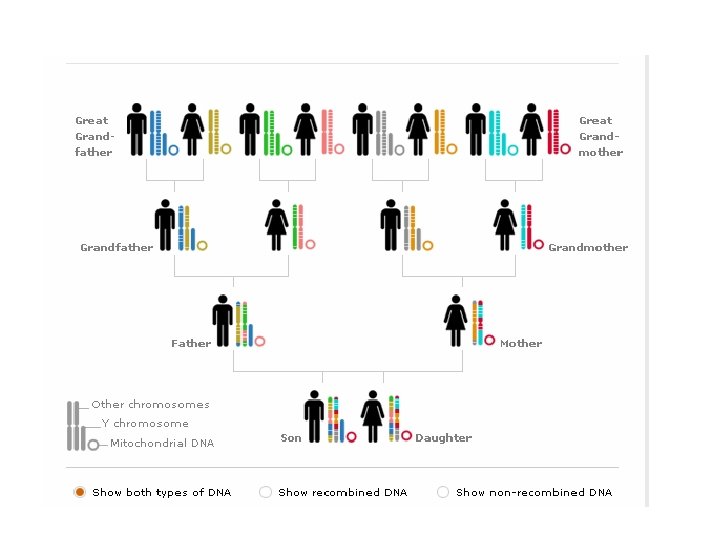

Kada su dva gena vezana na istom hromozomu, proporcija rekombinantnih gameta je manja od 50%

KROSING OVER se dešava na stupnju 4 hromatide

KROSING-OVER se dešava na stupnju 4 hromatide odnosno, posle replikacije hromozoma

višestruki C. O

Dva rekombinaciona dogadjaja koja se dese u istom regionu mogu poništiti medjusobne efekte; dogode se, ali ne uoče se Na većim distancama se mogu maskirati CO događaji preko dvostrukih CO.

RF – učestalost rekombinacije RF=broj rekombinanata/ukupan broj potomaka x 100 • Dvostruki CO, koji uključuje različite hromatide, može voditi različitim ishodima: • 0 rekombinanata • 50% rekombinanata • 100% rekombinanata

Hromozomske mape Trostruki heterozigot 3 vezana gena, dvostruki-krosing over Dvostruki CO među istim hromatidama daje 2 najmanje zastupljena rekombinantna fenotipa

relativne razdaljine i povezanosti gena u odnosu na učestalost rekombinacije razlika izmedju učestalosti rekombinacije (RF) i razdaljine izmedju gena. m. u. mere koliko se rekombinacije stvarno desilo izmedju gena, a RF govori o tome koliko je rekombinacija uočeno. RF=(SCO I + SCO II + DCO)x 100/total
 mape • Vezanost gena na hromozomu može se predstaviti genetičkom mapom -")
Hromozomske (genetičke) mape • Vezanost gena na hromozomu može se predstaviti genetičkom mapom - linearan raspored gena duž hromozoma • razmak između gena proporcionalan učestalošću rekombinacije izmedju njih • Jedinica razdaljine izmedju gena je c. M (centi Morgan) ili m. u (map unit), pri čemu 1 m. u. =1% rekombinacije Fizičke mape predstavljaju potpunu sekvencu DNK hromozoma sa razdaljinama i lokacijama gena merena brojem nukleotida

• Mapiranje hromozoma • Ukrštanje trostrukog heterozigota sa trostrukim recesivnim homozigotom • %rekombinanata . . . sledece. . .

Dvostruki CO među istim hromatidama daje 2 najmanje zastupljena rekombinantna fenotipa

Da li rekombinacioni dogadjaji na hromozomu utiču medjusobno? • Interferenca - CO u jednom regionu redukuje CO u susednom • dobijena učestalost DCO < očekivane učestalosti DCO (SCO I x SCO II) • Interferenca se računa preko koeficijenta koencidencije (C) • C=DCOdob/DCOoč • I =1 -C = 1 - DCOdob/DCOoč DCOdob=0, I=1 DCOdob< DCOoč, C -, I + geni su bliži DCOdob> DCOoč, C+, I - geni su udaljeniji

mapirajuća funkcija poboljšava tačnost RF u odnosu na rastojanje • • • 0, 1, 2, …i događaja m prosečan broj dogadjaja (srednji broj CO po mejozi) učestalost i-te klase: (i)=e-mmi/i! učestalost za i=0 je: (0)=e-mm 0/0!=e-m RF je m/2, polovina proizvoda onih mejoza koje su imale najmanje jedan CO u regionu: RF = 1/2(1 - e-m ) mapirajuća funkcija, za korekciju RF • što je m veće, e-m je manje, a RF dostiže max. 50%

• primer: • • na osnovu ukrštanja dobijeni RF=30%. Koliko je to u stvari m. u. , radaljine? 0. 30=1/2(1 -e-m) 0. 60=1 -e-m = 1 -0. 60=0. 40 iz tablice, za e-m =0. 40, korigovano m je 0. 90, RF korigovano je 45 m. u. korekcija razdaljine: nije 30 m. u već 45 m. u.

Analiza pojedinačnih mejoza i rekombinacije kod haploida • svi produkti mejoze dostupni na jednom mestu Uredjene i neuredjene tetrade

jedan hromozomski set, nema maskiranja ekspresije gena, fenotip je direktni odraz genotipa

Nema C. O. između centromere i lokusa Aleli A i a segregiraju u odvojena jedra posle I mejotičke deobe Segregacija M’ tipa • produkti mejoze su različiti zavisno od toga da li ima ili ne CO izmedju centromere i heterozigotnog lokusa u 2 n mejociti Ima C. O. između centromere i lokusa Aleli A i a segregiraju u odvojena jedra tek posle II mejotičke deobe Segregacija M’’ tipa

Zavisno od toga kako se centromere “prikače” na deobno vreteno u II mejotičkoj deobi razlikovaće se raspored spora u linearnom askusu

iz ukrštanja a+b+ x ab, svaki produkt je jedna haploidna jedinka • • • tipovi tetrada PD - parentalni ditip sadrže 2 parentalna genotipa NPD - neparentalni ditip - sadrže 2 neparentalna genotipa T -tetratip, sadrže 4 genotipa -mogu se dobiti bilo da su lokusi na odvojenim ili istim hromozomima ako je PD >NPD, lokusi su vezani ako je PD =NPD onda su na različitim hromozomima. Samo T i NPD sadrže rekombinante; NPD samo rekombinante, a T polovinu spora kao rekombinante RF = 1/2 T + NPD/ NPD+PD+T (x 100 u %). RF < 50%, lokusi su vezani
. • Ako")
Mapiranje centromere • primer: Neurospora crassa lokus za tip parenja (A, a). • Ako nema CO izmedju centromere i lokusa, spore A su na jednoj, spore a na drugoj strani askusa (M' tip segregacije). • Ako ima CO, izmedju centromere i lokusa, promena redosleda spora u askusu zavisno od toga da li imaju a ili A alel (M” tip segregacije). • Razdaljina lokusa od centromere = %rekombinanata, tj. tetrada koje pokazuju segregaciju M’’ /2.

Mitotičke rekombinacije • varijegacija, mozaicizam, stvaranje sektora, grupa ćelija u tkivu, koje imaju različit fenotip od tipičnog za to tkivo • Redak događaj, oko 1000 puta redje nego u mejozi • kod nekih gljiva, kvasaca, Drosophila • Povećana učestalost somatske mitotičke rekombinacije zapažena je nakon delovanja pojedinim mutagenima

Mitotičke rekombinacije • takodje na stupnju 4 hromatide • dovodi u homozigotno stanje lokuse distalno od CO (deo izmedju CO i kraja hromozoma) i na istom hromozomu, dalje od centromere.

Mapiranje genoma čoveka

Mapiranje genoma čoveka • Nisu moguća test ukrštanja • Moguća je: analiza rodoslova, upotreba molekularnih DNK markera –rekombinacione analize • razvojem molekularnih tehnika moguće je analizirati haploidne produkte mejoze razblažene do jedne ćelije, jednog spermatozoida, amplifikovane • detekcijom varijabilnih mesta na DNK kod neke osobe i njegovih haploidnih spermatozoida kao produkata mejoze, moguće je računati RF

X – hromozom prvi mapirani hromozom čoveka i to samo na osnovu rekombinacionih analiza

Korišćenje rekombinacija za mapiranja muški pol je hemizigot za X – dovoljno analizirati muško potomstvo od odredjene ženke, heterozigota za 2 vezana lokusa • Primer: • Muškarac sa retkim Xvezanim recesivnim poremećajem za metabolizam šećera (g) i slepilom za boje (c) daje potomke sa ženom bez ovih mutacija (CG/CG) • Ćerke su sve heterozigoti, sa dvostruko spregnutim CG/cg. • muško potomstvo ovih ćerki može dati uvid u RF - javljaju se parentalni i rekombinantni fenotipovi.

• Da bi se uoćili rekombinanti u potomstvu potrebne su "informativne mejoze„: veliki rodoslovi i potomstvo kod koga možemo, praćenjem pojave dve osobine za koje pretpostavljamo da se zajedno vezano nasledjuju, prepoznati rekombinaciju, tj pojavu nastanka osobina odvojeno • "NEINFORMATIVNA MEJOZA„ • A 1 B 1 • A 2 B 1 i rekombinantni i nerekombinantni gameti su isti A 1 B 1 ili A 2 B 1 • "INFORMATIVNA MEJOZA„ A 1 B 1 rekombinantni gameti su: A 1 A 2 B 2 nerekombinanti gameti su: A 1 B 1 A 2 B 2

DNK polimorfizam – varijabilnost u restrikcionim mestima i mapiranje hromozoma • Jedinka A je heterozigot za odredjeno restrikciono mesto Jedinka A • Jedinka B je homozigot za dugi fragment Jedinka B • Jedinke A i B mogu u medjusobnom ukrštanju dati deo potomstva sa 3 fragmenta, a drugi samo sa jednim.
 kao MARKER za mapiranje")
• • varijabilnost u restrikcionim mestima na DNK (RFLP) kao MARKER za mapiranje gena Ako je neko heterozigotno restrikciono mesto vezano za neki gen D u heterozigotnom stanju, u spregnutoj konformaciji, krosing over izmedju ova dva mesta daće rekombinantno potomstvo kao D-3 i d-2 -1. • RF je moguće izračunati izmedju različitih marker lokusa i izmedju marker lokusa i gena sa poznatim fenotipskim efektom.

• • Primer: Muškarac heterozigot za gen koji nosi neko oboljenje, D/d i za molekularni marker, M 1/M 2, žena homozigot za oba. Pedigre na slici daje genotipove dece u odnosu na gamete koji dolaze od oca. Dvoje su rekombinanti, RF = 33%

mape vezanosti gena • "LOD skor" izražava vezanost markera i gena • poredi verovatnoću da će odredjeni rezultati rodoslova da se dobiju ukoliko su 2 lokusa vezana i ukoliko nisu vezana • LOD skor je log 10 odnosa te dve verovatnoće • LOD skor >ili = +3, ukazuje na vezanost (log 101000=3) • Razdaljina medju lokusima se računa iz RFs koje daju najviši LOD skor • U humanom genomu, 1 m. u. odgovara 1 Mb.
 duž regiona hromozoma koji se nasledjuju zajedno")
Haplotip je set DNK sekvenci (SNP alela) duž regiona hromozoma koji se nasledjuju zajedno u haploidnom gametu x x x x x 5 različitih haplotipova 4 segregirajuća mesta rekombinacija u mejozi kod 1 roditelja izmedju 2 mesta daje rekombinantni haplotip

pregled • Geni na istom hromozomu su vezani i nasleđuju se zajedno • Ukrštanja koja uključuju vezane gene daju više zastupljenu parentalnu i manje zastupljenu rekombinantnu klasu, usled krosingovera, tj. mejotičke intrahromozomske rekombinacije • Krosing-over je recipročna razmena delova homologih hromozoma između nesestrinskih hromatida za vreme profaze I mejoze • Što su geni udaljeniji na hromozomu, veća je šansa za rekombinaciju, ali i za višestruke krosing-overe • Mape vezanosti zasnivaju se na odnosu krosing overa i rastojanja među genima i mapirajuće funkcije se koriste da isprave previd učestalosti dobijenih rekombinacija • Analiza tetrada je tehnika mapiranja gena kod haploidnih eukariota, kod kojih produkti pojedinačne mejoze ostaju zajedno u askusima • Rastojanje među genima analizom tetrada određuje se relativnom porporcijom tipova tetrada • Krosing over se može dešavati i u mitozi i tada nastaje mozaični fenotip ili se stvaraju različiti klonovi ćelija • Za prepoznavanje rekombinanata i mapiranje gena u humanom genomu koriste se rodoslovi i molekularni markeri, vezani za lokuse koji odredjuju neku osobinu, u heteozigotnom stanju.
- Slides: 41