MHC genes and Products Chromosome 6 contains human










\" determine which")






 compares the properties of MHC class I and")













- Slides: 38
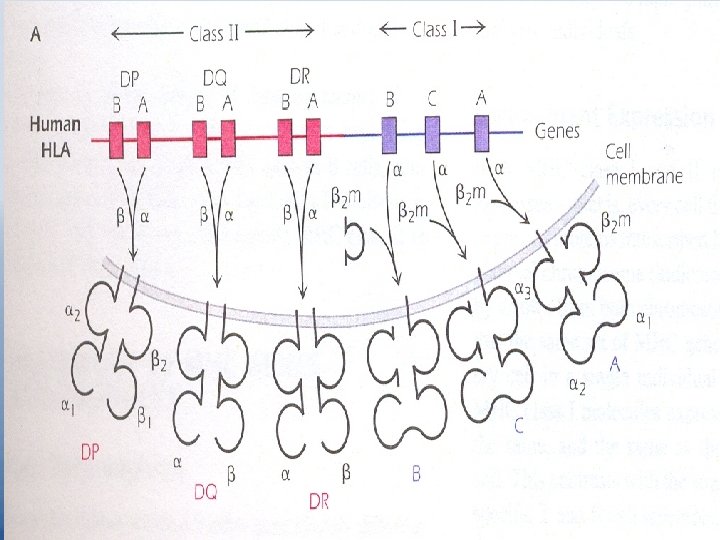
MHC genes and Products – Chromosome 6 contains human MHC called HLA "Human Leukocyte Ag". – Two sets of MHC genes MHC class II And their cell – surface expressed products "MHC Ags". – Fig 9. 1 Page 114. 3 independent genes code for MHC class I molecules they are HLA – A HLA – B HLA – C Human MHC class II region is made up of 3 sets of genes HLA – DP HLA – DQ HLA – DR
– Each MHC class II subregion contains an A and a B gene that codes for one chain, or respectively, of a two-chain MHC class II molecule. E. g. HLA-DPA gene codes for DP of the HLA-DP molecule and the HLA-DPB gene codes the other chain DP , of the HLA -DP molecule. The two chain molecules DQ and DR are made by similar pairing of gene products of the DQ and DR subregions. – MHC class I molecules are expressed on every nucleated cell. – MHC class II molecules have more limited distribution on B cells, dendritic cells, thymic epithelial cells, Macrophages and monocytes. "APCs".
– Expression of MHC class I is coordinate, i. e. all 3 MHC class I molecules are expressed on the cell surface at the same time. – MHC class II are also coordinately expressed, but under distinct regulation. – MHC class I can be expressed in the absence of any MHC class II. – The level of MHC class I & II expression at the cell surface can be coordinately upregulated or down regulated by a number of stimuli, e. g. IFN enhances expression of all MHC class I & II.
Variability of MHC genes and Products Genetic Polymorphism Different individuals within a species have slightly different forms, referred to as alleles, of each MHC class I or II gene. At a single MHC locus, different individuals have different types of prototypical gene. In humans, different alleles are given numbers such as: HLA-B 15 HLA-B 27 The phenomenon of having multiple stable forms of one gene in the population is known as genetic Polymorphism which generates diversity of MHC within the population.
– MHC is the most highly polymorphic gene system in the body and hence in the population. – This extensive polymorphism of MHC genes makes it very unlikely that two random individuals will express identical sets of MHC molecules. – This polymorphism is the basis of rapid graft rejection between genetically different individuals.
Codominant Expression – Both MHC class I and II molecules are codominantly expressed "from both the maternal and paternal chromosome". – Every cell within one person expresses up to 6 different HLA class I molecules, 3 coded for by HLA class genes on each chromosome. e. g. all cells in one person may express the following molecules on their surface: HLA-A 2 HLA-A 5 HLA-B 7 HLA-B 13 HLA-C 6 HLA-C 8 Another person may express 6 completely different HLA class I molecules.
– Some individuals may express fewer than 6 different class I molecules because they are homozygous for a particular allele "both paternal and maternal chromosomes express the same allele". – HLA DP, DQ and DR molecules are also expressed codominantly in cells expressing HLA class II molecules. Frequently more than 6 molecules are expressed, because gene products from different genetic region – such as DP with DQ - can pair and be expressed at cell surface.
– The set of MHC I and II genes expressed on an individual chromosome. – Referred to as a haplotype – is passed on as a unit into the offspring. – HLA haplotypes of any offspring will differ from the haplotypes of parents. – MHC identity occurs in monozygotic twins and can occur in a family of large number of children. – It is unlikely to find individuals in the general population who are matched at all HLA alleles.
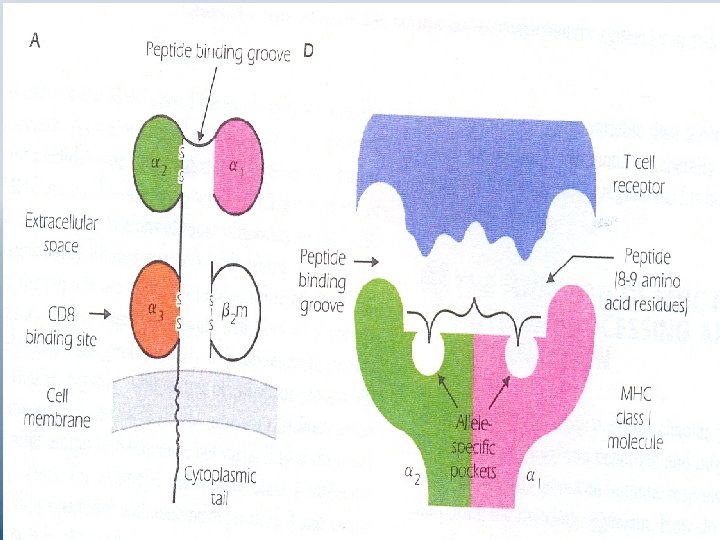
Structure of MHC Molecules - MHC class I molecules Fig 9. 3 p 117 – Each MHC class I gene codes for a transmembrane glycoprotein of approximate molecular weight 43 KDa. which is referred to as or heavy chain. – It comprises 3 extracellular domains 1, 2 and 3. – 2 -microglobulin is associated non-covalently with MHC class I molecule. 2 m is coded for on another chromosome. It resembles a single Ig domain. It is a member of Ig super family.
– At cell surface MHC class I plus 2 m has the appearance of a four-domain molecule. – Sequence differences among MHC molecules are confined to a limited region in their extracellular 1 and 2 domains. – An individual class I molecule can be divided into a non-polymorphic or invariant region which is similar in all class I allelic forms, and a polymorphic or variable region "sequence unique to that allele". CD 8 binds to the invariant region of all MHC class I molecules.
– Allele-specific pockets in the peptide binding groove "Fig 9. 3 (D)" determine which peptides bind to a particular allelic form of and MHC molecule. the pockets also help secure peptides in a position in which they can be recognized by specific TCRs. – One MHC molecule can bind to a large number of peptides with different sequences.
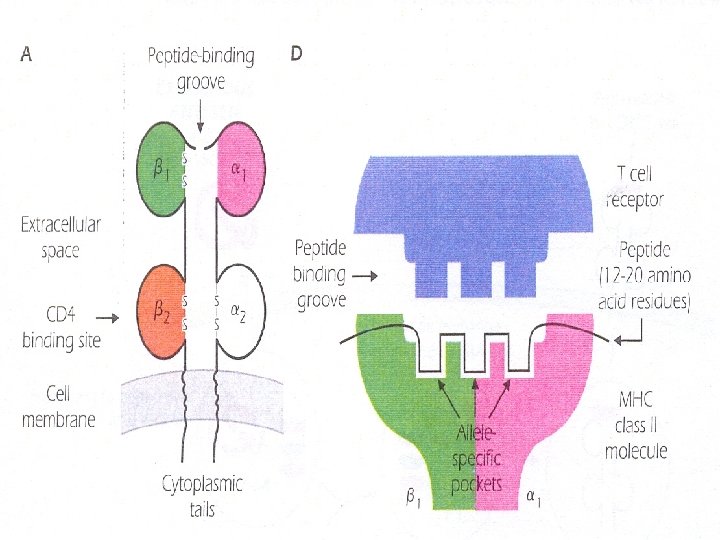
Structure of MCH class II Molecules – MHC class II and code for chains of approximate M. wt. – Like class I, they are transmembrane glycoprotein molecules with cytoplasmic tails and extracellular Iglike domains " 1, 2, 1, 2". – MHC class II molecules are members of Ig superfamily. – MHC class II molecules are also made up of variable or polymorphic regions "differing among alleles" and invariant or nonpolymorphic regions "common to alleles".
§ The T cell molecule CD 4 binds to the invariant portion of all MHC class II molecules. § MHC class II molecule also contains peptide-binding groove or cleft at the top of the molecule which is structurally analogous to the MHC class I groove. § Exogenous protein Ags are taken into APCs and processed to peptides in acid compartments where they interact with MHC class II molecules. § The peptide-MHC class II complex is transported to the cell surface where it interacts with the TCR expressed by a CD 4+ T cell.
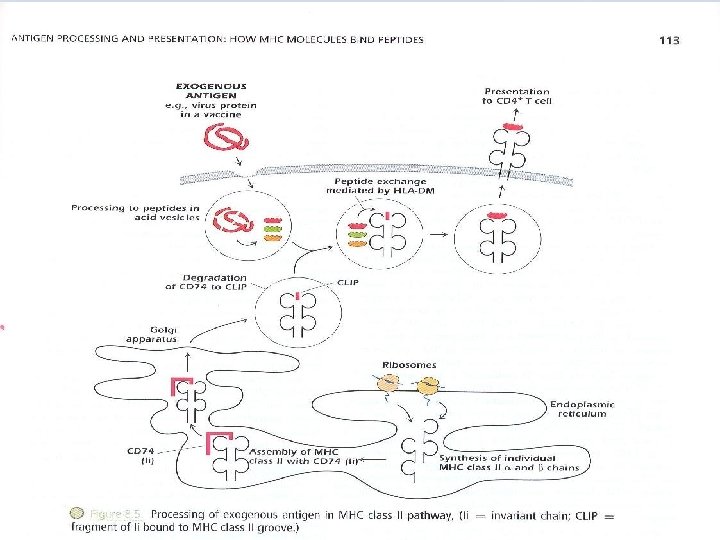
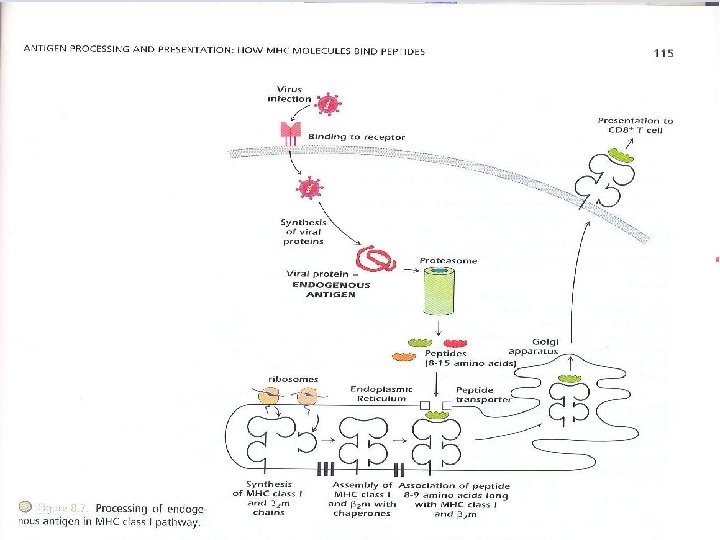
– The response of CD 4+ T cells is referred to as restricted by MHC class II molecules – "self-MHC class II molecules". – Exogenous Ags can be derived from pathogens (such as bacteria and viruses) or from foreign proteins( such as vaccines) that do not injure the host but activate an immune response. – Fig 8. 5 shows the processing and presentation of a typical exogenous Ag, a protein is injected as a component of inactivated or “dead” virus vaccine. – The protein is internalized, contained in an intracellular vesicle that fuses with endosomal or lysosomal vesicles that are highly acidic.
– These vesicles contain an array of proteolytic enzymes , proteases known as cathepsins, which function at low p. H, cut proteins into peptides. The MHC class II α and β chains are synthesized individually in ER and are assembled there with CD 74, invariant chain (Ii) CD 74 binds to the groove of MHC class II molecule preventing other peptides from binding CD 74 also acts as chaperone i. e MHC leaves ER to Golgi complex; from there, they proceed into the acid vesicle endocytic pathway.
Removal of CD 74 occurs in stages; initially it is degraded leaving a fragment known as CLIP(class II – associated invariant polypeptide) bound to MHC class II groove. Vesicles containing MHC class II bound to CLIP then fuse with acid vesicles containing peptides derived from the catabolism of exogenous Ags. In this compartment, a molecule known as HLA-DM catalyzes peptide exchange between the MHC class II-CLIP complex and peptides derived from the exogenous Ags. – Endogenous protein Ags, generally derived from infectious pathogens, come from within cells and are processed to peptides in the cytosol. Peptides derived from these proteins interact with MHC class I molecules in the ER.
– The peptide MHC class I complex is transported to the cell surface where it interacts with the T cell receptor expressed by a CD 8+ T cell. The response of CD 8+ T cell is referred to as restricted by MHC class I molecules "self-MHC class I molecules". – Because MHC class I molecules are expressed on all nucleated cells, the processing and presentation of endogenous Ags can occur in every cell in the body. – Because pathogens can infect any cell in the body, CD 8+ T cells can scan MHC class I and peptide combinations expressed on any nucleated host cell to identify whether it has been infected.
– Table 9. 1 (p. 141) compares the properties of MHC class I and class II molecules. – Proteins from bacteria, viruses, allergens or harmless Ags, trigger CD 4+ T cell response. – Pathogens infecting cells "bacteria, viruses" create epitopes that are presented to CD 8+ T cell. – The processing pathway, rather than inherent properties of Ag determines whether a protein is presented to CD 4+ or CD 8+ T cell, this is illustrated by the – Cross presentation pathway in which an exogenous Ag ends up being – Processed in the MHC class 1 pathway and presented to CD 8+ T cells.
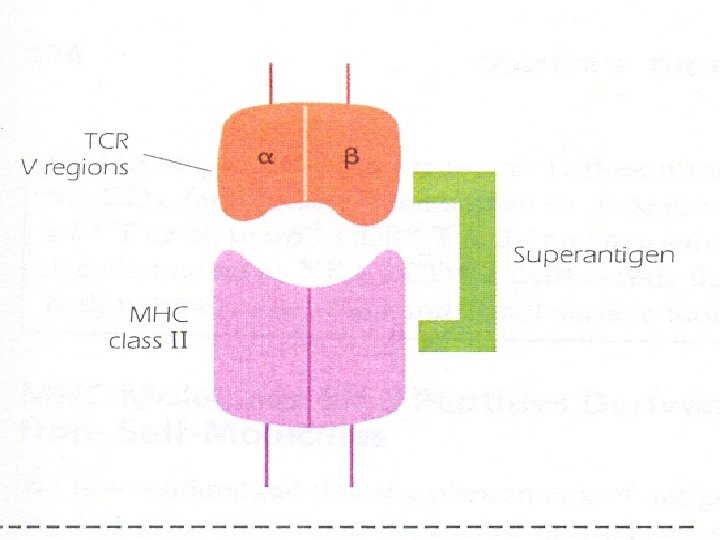
– Superantigens: – They are bacterial toxins such as S. aureus enterotoxin. Superantigens bind to MHC class II molecules without being processed. Superantigens bind to a region of the MHC class II molecule outside the peptide-binding groove and bind as well to the V region of TCR. – Superantigen activates a huge number of an individuals T cells "up to 10 % of total number" leading to their apoptosis. – Activation of many T cells can have clinical consequences; for example, it is – Responsible for inducing the shock-like syndrome of toxic shock.
Inability to respond to an Ag: – Costimulators are induced by pathogens in cells presenting Ag to T cells. Costimulator signals activate naïve T cells "cells have not previously encountered Ag". – Peptides derived from self-molecules generally encounter T cells on normal tissue cells "e. g. of liver and pancreas" which do not express costimulator function, in this case, T cells are not activated, even if peptides derived from a self-molecule are presented by APC in the tissue cell. T-cells are not activated because APCs in tissue do not normally express co-stimulator signals in the absence foreign Ag or of an inflammatory response.
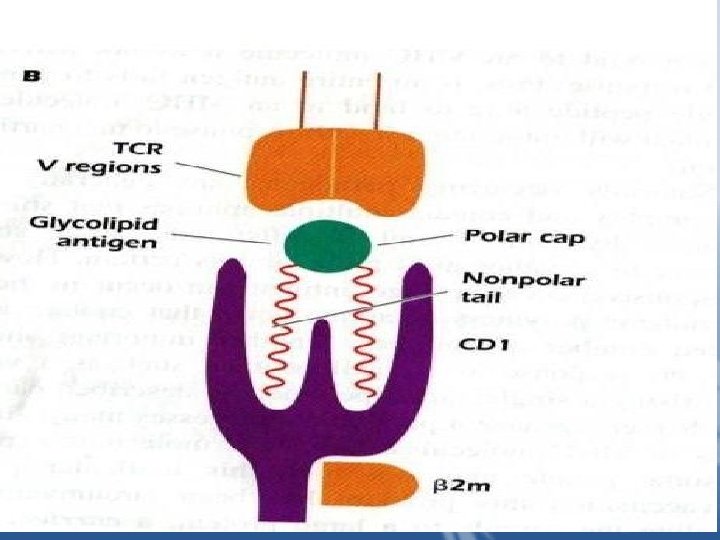
– Exposure to foreign Ag induces tissue damage "the danger hypothesis" and that the danger signals activate T cells. This hypothesis explains why T cells respond to foreign Ag but not to self molecules. Lipids and Glycolipids presented by CD 1 To NKT Cells Some T cells can recognize lipids and glycolipids found in the cell walls of pathogens Such as Mycobacterium tuberculosis that live inside macrophages. These molecules Are presented by CD 1(CD 1 a-CD 1 d) expressed by APC such as macrophages and dendritic cells.
CD 1 molecules are cell surface glycoproteins expressed on the surface of APC in Association with β 2 m. The structure of CD 1 molecule is similar to that of an MHC class 1 molecule, but CD 1 contains a larger binding groove with a deep cavity. The Cavity binds the hydrophobic backbone of a lipid Ag exposing the polar region of a Lipid for binding TCR. Binding of lipid Ag to CD 1 is believed to take place in acidic cellular compartments, Similar to loading of exogenous peptides onto MHC class 11 molecules. CD 1 a , b and c present lipids to T cells that use an αβ TCR, but CD 1 d present lipids and glycolipids to NKT cells.
MHC and Immune Response 1. MHC molecules play a crucial role in the response of T cells to antigens that penetrate or live inside cells of the body. MHC molecules bind peptides derived from proteins and present them to T cells with the appropriate receptor. Thus T cell responses are said to be MHC restricted. 2. The MHC codes for two major categories of cell-surface transmembrane molecules, MHC class I and class II molecules. MHC class I molecules are expressed on all nucleated cells; MHC class II molecules are expressed constitutively on APCs such as B cells and dendritic cells. The expression of MHC molecules is inducible on many cell types, particularly in response to cytokines released during the response to infectious agents.
3. Each individual expresses a distinct array of MHC class I and class II molecules. This diversity comes about because different individuals within a species have a range of slightly different forms (alleles) of MHC class I and class II genes (genetic polymorphism). Because of the extensive polymorphism of MHC genes, every individual has an almost unique array of inherited MHC genes. 4. Within one individual, the MHC class I and class II molecules expressed are the same on all cells of the body, and they are codominantly expressed at the cell surface (i. e. , products of both maternal and paternal chromosomes).
5. The outer region of every MHC class I and class II molecule contains a deep groove that binds peptide derived from the catabolism (processing) of protein antigens. The binding of peptides to MHC molecules is selective. Each MHC molecule binds a distinct subset of peptide with a particular motif. 6. Exogenous protein antigens are taken into APCs and processed to peptides in acid compartments where they interact with MHC class II molecules. The peptide. MHC class II complex is transported to the cell surface where it interacts with the TCR expressed by a CD 4+ T cell. The response of CD 4+ T cells is referred to as restricted by MHC class II molecules (self-MHC class II molecules).
7. Endogenous protein antigens, generally derived from infectious pathogens, come from within cells and are processed to peptides in the cytosol. Peptides derived from these proteins interact with MHC class I molecules in the endoplasmic reticulum. The peptide-MHC class I complex is transported to the cell surface where it interacts with the T cell receptor expressed by a CD 8+ T cell. The response of CD 8+ T cells is referred to as restricted by MHC class I molecules (self-MHC class I molecules). 8. Since proteins are generally structurally complex, they usually generate at least one peptide able to bind to an MHC molecule, ensuring that a T cell response is made to at least some part of a foreign antigen.
9. MHC molecules bind peptides derived from selfcomponents as well as from foreign antigens, but complexes of MHC molecules with self-peptides do not normally activate a T cell response. This is because self-molecules do not normally generate the costimulatory (second) signals needed to activate naïve T cells. The T-cell response is focused on the response to foreign (non-self) molecules, particularly components of microorganisms, which induce costimulatory function. 10. Susceptibility and resistance to many diseases in humans and other species are associated with the expression of a particular MHC allele.
11. Cytokines synthesized in response to infectious agents induce or increase the expression of MHC class I and class II molecules. This leads to enhanced immune response to the pathogen that induced the response. In contrast, some viruses ( such as the herpes simplex virus, adenovirus, and cytomegalovirus) synthesize proteins that interfere with processing of endogenous Ag pathway; they inhibit the synthesis of MHC class I molecules or interrupt the transport of peptide-MHC class I complexes to the cell surface. In this way the virus decreases expression of MHC class I molecules and so decrease the CD 8+ T-cell response to the virus. 12. Tumor cells frequently show decreased expression of MHC class I compared to normal cells, subverting a potential CD 8+ T-cell response.
13. Although the decrease in MHC class I expression decrease the CD 8+ T-cell response, it may also act as a trigger of NK-cell responses to the virus-infected cell or tumor cell. Cross presentation: Exogenous Ags presented in the MHC class I pathway APC and particularly dendritic cells have a unique pathway, called cross presentation, for generating peptides derived from exogenous protein Ags and presenting them to CD 8+ T cells. This pathway is important in activating CD 8+ T cells to respond to tissue cells infected by some viruses that are not taken up by APC as well as response to dying cells.
Bare Lymphocyte Syndrome People lacking the ability to express either HLA class I or class II molecules or both. Individuals who do not express class II molecules have immunodeficiency charachterized by defective presentation of Ags to CD 4+ T cells and decreased numbers of CD 4+ T cells. These individuals have a mutation in one of the factors that controls the transcription of MHC class II genes.