SNA Caratteristiche comuni e specifiche Funzioni fisiologiche Attivazione























- Slides: 27
SNA Caratteristiche comuni e specifiche Funzioni fisiologiche Attivazione Sne Controllo centrale e interazioni con il sistema neuroendocrino Nel 1889 John Langley domostrò che dai gangli fuoriuscivano assoni di neuroni che si potevano eccitare con la nicotina e questi neuroni attivati producevano nell’organismo intense risposte periferiche come vasocostrizione, sudorazione, piloerezione. Successivamente in Australia John Eccles e la figlia Rosamund studiarono i potenziali sinaptici nei gangli simpatici scoprendo fenomeni interessanti. Nei primi anni del novecento, Walter Cannon delineò il significato funzionale del SNA collegato all’omeostasi. Il SNA mette in equilibrio l’ambiente interno bilanciando l’ attivazione del simpatico, del parasimpatico, enterico con una funzione fondamentale svolta dall’ipotalamo
Al SNA appartiene anche il sistema nervoso enterico localizzato nell’apparato gastrointestinale Neuroni pregangliari sono localizzati nei nuclei del tronco dell’encefalo per il parasimpatico e nel midollo per simpatico e parasimpatico. Questi sono neuroni mielinizzati con velocità di conduzione pari a 2 -5 m/s dirette ai gangli. I gangli sono stazioni di controllo, ricevono non solo neuroni pregangliari, ma anche fibre nervose sensoriali dagli organi addominali che formano circuiti riflessi coinvolti nel controllo di alcune funzioni viscerali. NB i gangli parasimpatici ricevono anche fibre noradrenergiche dai gangli simpatici.
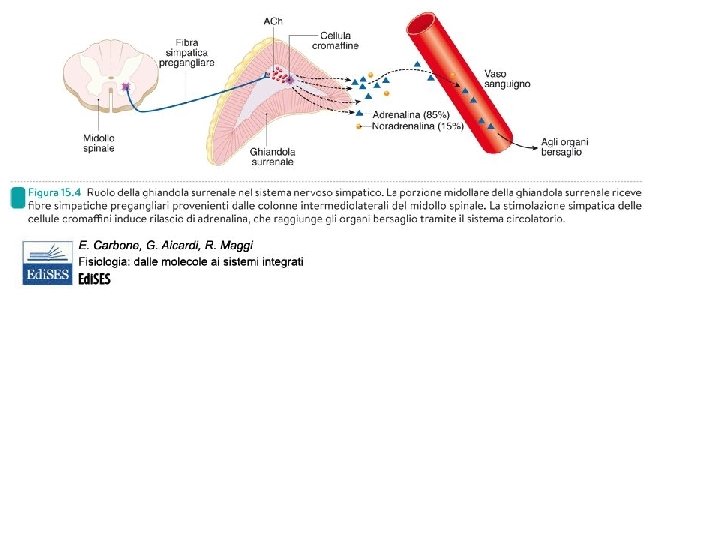
Neuroni simpatici pregangliare si trovano delle colonne intermedio-laterali della sostanza grigia spinale dal primo segmento toracico al quarto lombare. Gli assoni (mielinici) escono dalle radici ventrali insieme ai nervi somatici spinali (nervi spinali) da cui si separano formando i rami comunicanti bianchi Alcuni assoni simpatici decorrono con i nervi splancnici diretti alla midollare del surrene
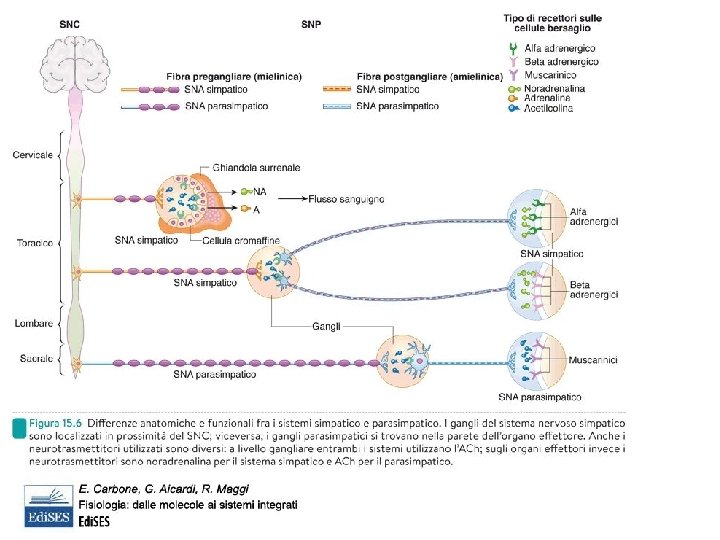
I gangli simpatici sono ai lati della colonna nei gangli paravertebrali oltre al ganglio celiaco mesenterico superiore e inferiore detti prevertebrali. La fibra pregangliare può fermarsi e sinaptare direttamente nel ganglio o percorrere la catena paravertebrale verso l’alto o verso il basso e sinaptare a distanza. La fibre postgangliari (amieliniche) ritornano ai nervi spinali attraverso i rami comunicanti grigi e arrivano a destinazione. La Acetilcolina attiva i recettori nicotinici gangliari dando un potenziale postsinaptico eccitatorio rapido della durata di 10 -20 msec in grado di attivare un potenziale d’azione. L’Acetilcolina attiva anche recettori muscarinici gangliari dando un potenziale postsinaptico eccitatorio lento della durata di 0. 5 -1 sec sia inibitorio che eccitatorio con la funzione di modulare la trasmissione gangliare rapida. Le fibre sensitive rilasciano nel ganglio peptidi come sostanza P, peptide correlato al gene della calcitonina (CGRP) che danno potenziali eccitatori o inibitori della durata dell’ordine del minuto, con la funzione di modulare la trasmissione gangliare rapida. Nei gangli troviamo anche i neuroni intrinseci, localizzati interamente nel ganglio, che rilasciano le catecolamine dopamina e noradrenalina, serotonina, istamina, sostanza P, encefaline, e somatostatina. Questi modulano la trasmissione gangliare rapida.
Nel ganglio ci sono i neuroni postgangliari che ricevono contatti dai neuroni pregangliari. I neuroni pregangliari fanno sinapsi anche con numerosi neuroni postgangliari (DIVERGENZA). Nei gangli simpatici la fibra pregangliare prende contatto con 9 neuroni postgangliari andando a influenzare ben 9 siti diversi (attivazione “lotta o fuga”). Nei gangli parasimpatici la divergenza è minore (rapporto 1: 3 o anche 1: 1). Nei gangli si verifica anche la CONVERGENZA, un neurone postgangliare riceve numerse fibre pregangliari da neuroni diversi. Questo rende possibile la sommazione spaziale di potenziali postsinaptici e l’integrazione.
Neuroni da organi periferici Neuroni dal SN centrale Neuroni da altri gangli Convergenza o divergenza segnali Trasmissione rapida Modulazione da trasmissione lenta Potenziamento o depressione
La trasmissione colinergica gangliare va incontro a plasticità sinaptica come potenziamento e depressione a lungo termine, rispettivamente un incremento o diminuzione della trasmissione sinaptica di durata variabile da ore a giorni. Dal ganglio parte il neurone postgangliare amielinico a bassa velocità di conduzione (0. 5 -2 m/s) diretti ai muscoli lisci degli organi, dei vasi delle vie respiratorie, al muscolo cardiaco, l’occhio, le ghiandole esocrine ed endocrine e il tessuto adiposo: giunzione neuroeffettrice Ach + peptidi neuromodulatori NA + peptidi neuromodulatori
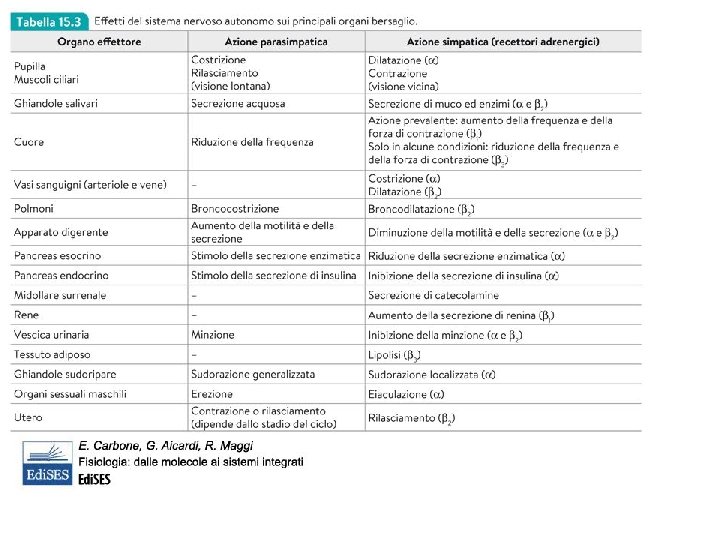
lipolisi Effetto ionotropo e cronotropo negativo, vasodilatazione Alpha 1 e beta 2 sono localizzati anche nella parete del tubo digerente e nelle ghiandole esocrine sui quali inducono effetti opposti (rispettivamente contrazione e rilassamento). Alpha 2 si trovano solo sulla muscolatura liscia di stomaco e intestino (e arteriole cutanee e coronariche). DESTINO DEI NEUROTRASMETTITORI: NA viene ricaptata dal terminale neuronale e reimmagazzinata nelle vescicole. Una piccola parte viene ossidata dalle MAO, monoaminoossidasi mitocondriali all’interno dei neuroni o della glia ma NON nel sangue. NA e A circolanti vengono inattivate per metilazione nel fegato e nel rene da COMT, catecol. O-metil-transferasi
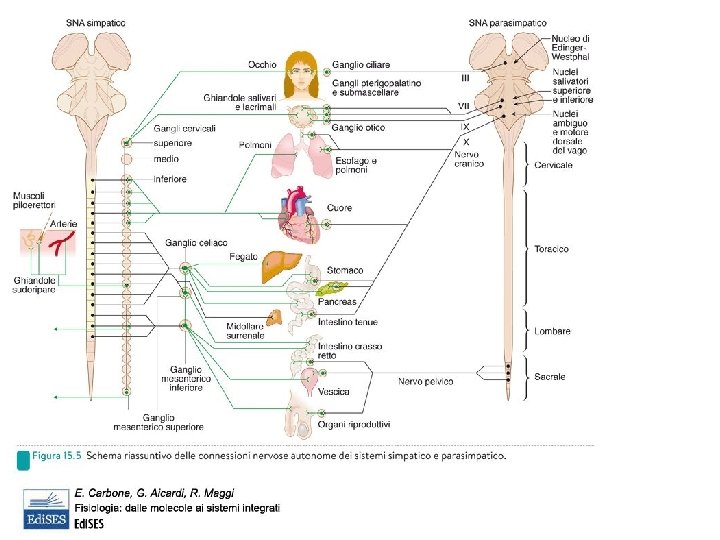
PARASIMPATICO I neuroni parasimpatici pregangliari sono localizzati nei nuclei di alcuni nervi cranici e nei segmenti S 2 -S 4 del midollo spinale. Dal nucleo di Edinger-Westphal nel mesencefalo originano parasimpatiche pregangliari che decorrono nel nervo oculomotore (III paio nervi cranici) e terminano del ganglio ciliare da cui originano le fibre post-gangliari dirette all’occhio. Dal nucleo salivatorio superiore e inferiore situati nel ponte e nel bulbo originano parasimpatiche pregangliari che decorrono nel nervo facciale (VII paio nervi cranici) e glossofringeo (IX) che terminano nei gangli parasimpatici pterigopalatino, submascellare e otico da cui originano fibre post-gangliari per le ghiandole salivari. Dal nucleo motore dorsale del vago e del nucleo ambiguo, nel bulbo, originano la maggiorparte delle fibre parasimpatiche pregangliari che decorrono nel nervo vago (X paio) che terminano in plessi all’interno degli organi, plesso cardiaco, paratracheale, bronchiale e pancreatico) da cui originano le postgangliari per cuore, polmone, apparato gastrointestinale con fegato, stomaco, intestino tenue, primo tratto del crasso e pancreas). Dal segmento S 2 -S 4 originano pregangliari che decorrono nei nervi pelvici dirette ai gangli localizzati in parete distale dell’intestino, vescica e organi sessuali
ACETILCOLINA M 1 più espresso nel SNC, ghiandole esocrine e gangli simpatici M 2 cuore e muscolo liscio delle vie respiratorie, apparato gastrointestinale e urinario M 4 e M 5 nel SNC M 2 aumentano la peristalsi mediante apertura dei canali del sodio e del calcio DESTINO: degradazione mediante acetilcolinesterasi sia nel vallo sinaptico che nel sangue
FUNZIONI FISIOLOGICHE Tono basale rispetto al quale il simpatico e/o il parasimpatico incrementano o riducono l’attività. SIMPATICO: lotta e fuga. Effetti sistemici sull’intero organismo e lenti PARASIMPATICO: riposo e digestione. Effetti localizzati e veloci L’attivazione è riflessa e involontaria in risposta agli stimoli sensoriali: 1 -riflessi oculari come regolazione del diametro pupillare e messa a fuoco del cristallino 2 -riflessi cardiovascolari: gittata cardiaca, resistenza periferica 3 -riflessi ghiandolari: secrezioni nasali, lacrimali, sudoripare 4 -RIFLESSI GASTROINTESTINALI: controllo motilità e secrezioni dell’apparato digerente 5 - riflessi urogenitali: controllo minzione e funzioni sessuali LOTTA O FUGA: condizioni di stress o durante esercizio fisico: aumento gittata cardiaca, vasocostrizione periferica con aumento pressione arteriosa e vasodilatazione muscolare con aumento flusso ematico, broncodilatazione e aumento ventilazione polmonare, attivazione SNC, dilatazione pupillare, sudorazione, aumento glicemia … catabolismo e mobilizzazione energia. Contemporaneamente inibizione digestione, defecazione e minzione. Riposo e digestione: anabolismo e ripristino riserve energetiche
SISTEMA NERVOSO ENTERICO O PICCOLO CERVELLO Nella parete del tubo digerente ritroviamo il plesso mioenterico di Auerbach che si estende dall’esofago allo sfintere anale interno e il sottomucoso di Meissner che non è presente nell’esofago e nello stomaco. Nei plessi esistono circa 14 neuroni diversi: sensoriali, neuromotori, interneuroni NEUROTRASMETTITORI responsabili delle motilità 1 -serotonina e Ach stimolano la contrazione muscolatura liscia 2 -ATP, dopamina e NO inibiscono la muscolatura liscia 3 -Peptidi ad azione stimolatoria: colecistochinina, encefaline, peptide rilasciante la gastrina, neuropeptide Y, sostanza P 4 -Peptidi ad azione inibitoria: somatostatina, polipeptide intestinale vasoattivo VIP NEUROTRASMETTITORI responsabili della vasodilatazione-vasocostrizione e funzioni secretorie esocrine (succhi digestivi) ed endocrine (rilascio ormoni)
Riflessi intrinseci, brevi e lunghi
Riflesso estrinseco lungo: riflesso inibitorio intestino-intestinale Se una regione dell’intestino subisce una distensione intensa e duratura, i meccanocettori in via riflessa causano un rilasciamento della parte intestinale adiacente alla zona distesa. Resezione del nervo vago produce atonia gastrica e intestinale, riduzione drastica della motilità e costipazione. In condizioni di stress, rabbia o paura, con forte attivazione del simpatico, diminuisce la peristalsi e aumenta il tono degli sfinteri, e il cibo viene trattenuto nel canale alimentare Accoppiamento tra VIP e NO: NO causa il rilascio di VIP, VIP si lega ai recettori sulle cellule liscie e attiva la NOS che produce NO che retrodiffonde inducendo rilascio di VIP etc…. risulatato rilasciamento muscolatura liscia
CONTROLLO CENTRALE DEL SNA
SNA riceve afferenze da sistema limbico, nucleo del tratto solitario, corteccia e ipotalamo. Il nucleo del tratto solitario, bulbare, riceve informazioni dal nervo facciale, glossofaringeo e vago e le utilizza per modulare il tono vagale sul cuore e sull’apparato gastrointestinale. Il sistema limbico (amigdala, nucleo mammillare del talamo, ipotalamo, bulbo olfattivo e ippocampo) che conserva i ricordi di esperienze olfattive, visive ed uditive, gestisce sensazioni subcoscienti in grado di modulare il SNA. Stati emozionali intensi possono indurre risposte viscerali, come tachicardia, aumento pressione arteriosa, sudorazione. Corteccia cerebrale controlla volontariamente defecazione e minzione, dopo opportuno training può controllare anche lo stress psichico. L’ipotalamo, centro di controllo del sistema endocrino, ha un ruolo centrale nel controllo del SNA. Riceve informazioni sensitive viscerali, visive e olfattive, integra stimoli legati alla variazione della temperatura corporea, dell’osmolarità, della glicemia, della concentrazione del sodio. L’ipotalamo integra tutte queste informazioni afferenti dall’ambiente interno ed esterno, li confronta con i propri sensori ipotalamici (osmocettori, termocettori, recettori ormonali) e avvia risposte complesse involontarie e comportamentali
Il nucleo paraventricolare dell’ipotalamo è il principale centro di integrazione, qui ritroviamo neuroni pregangliari del SNA che regolano il flusso di informazioni verso il SNA simpatico e parasimpatico e neuroni ad attività neuroendocrina. Il mantenimento dell’omeostasi è garantito dal lavoro concertato del sistema autonomo e di quello neuroendocrino nel nucleo paraventricolare. IPOTALAMO E SNA: PRESSIONE ARTERIOSA (controllo tono vasomotore) BILANCIO IDROELETTROLITICO (controllo assunzione acqua e sale) TEMPERATURA CORPOREA (termogenesi e risposte comportamentali) METABOLISMO ENERGETICO (assunzione cibo, digestione, metabolismo) ATTIVITA’ RIPRODUTTIVA STRESS
Le risposte neuroendocrine, involontarie e comportamentali sono mediate da tre sistemi efferenti: 1 -segnali diretti all’eminenza mediana per il controllo della secrezione degli ormoni dell’adenoipofisi 2 -segnali diretti all’ipofisi posteriore per il controllo della secrezione degli ormoni liberati dalla neuroipofisi 3 -proiezioni discendenti ai centri simpatici e parasimpatici del mesencefalo e del midollo spinale (nucleo del tratto solitario, nucleo motore dorsale del vago, midollo intermedio laterale) 4 -proiezioni ascendenti alle regioni limbiche attivano centri corticali per le risposte comportamentali
Innervazione gastrointestinale
SIMPATICO: dal ganglio celiaco le fibre postgangliali innervano: stomaco, tenue, fegato, pancreas : dal ganglio ipogastrico al crasso PARASIMPATICO: fibre pregangliali provengono dal nucleo del Vago (esofago, stomaco, tenue, cieco, colon ascendente e metà colon trasverso) o dai nuclei sacrali S 2 -S 4 (il resto del crasso). Le fibre post-gangliali originano dai plessi intramurali di Meissner e di Auerbach, se vengono assimilati ai gangli parasimpatici PLESSI NERVOSI INTRINSECI di Meissner (sottomucoso) e di Auerbach (mioenterico) che comprendono neuroni sensoriali, interneuroni e neuroni motori e cellule interstiziali di Cajal
L’attività dell’apparato digerente, seppur regolata dal sistema nervoso autonomo, è controllato direttamente dai plessi di Auerbach (mioenterico) e Meisser (sottomucoso) che costituiscono la rete neuronale con: - Cellule gangliari cui afferiscono le fibre parasimpatiche - Fibre afferenti sensitive che provengono dalla mucosa - Interneuroni - Fibre efferenti direttte alle cellule secretorie e alle cellule muscolari lisce della parete intestinale - - - I pessi di Auerbach e Meissner danno origine al plesso enterico costituito dal plesso terziario che innerva la muscolatura longitudinale, il plesso muscolare profondo che innerva la muscolatura circolare e il plesso mucoso che innerva la mucosa. Il sistema enterico permette un controllo autonomo o semiautonomo dell’attività motoria e secretoria dell’apparato digerente, infatti la presenza di neuroni sensoriali, interneuroni e neuroni motori permette la realizzazione di riflessi locali. L’autoritmicità è a carico delle cellule interstiziali di Cajal, cellule intermedie tra le mesenchimali e le lisce che si trovano raggruppate nel plesso mioenterico di Auerbach o nello strato muscolare. Nell’esofago, nello stomaco e nel colon le Cajal si trovano nello strato circolare mentre nel tenue nello strato muscolaris muscolarae. Le cellule di Cajal presentano stretto contatto con le cellule autonome e giunzioni comunicanti con le cellule lisce così che l’autoritmicità si propaga alle cellule muscolari adiacenti.
Cellule di Cajal: ritmo elettrico di base con potenziali lenti, spontanei e autoritmici che si propagano alle cellule muscolari adiacenti: REB dello stomaco è di 3 cicli al minuto; REB duodeno 12 cicli al minuto. Nelle cellule lisce il REB è presente anche in assenza di contrazione che avviene solo quando il REB è soprasoglia, quando si manifestano potenziali d’azione, in genere sul plateau del REB e maggiore è la frequenza dei potenziali d’azione maggiore è il grado di contrazione, poiché maggiore è la concentrazione del calcio intracellulare. Inoltre lo strato ad anello si contrae più velocemente di quello longitudinale
Controllo nervoso della motilità intestinale Il SNA influenza la motilità locale e controlla attraverso riflessi intestinali lunghi la motilità di segmenti distanti. La stimolazione parasimpatica incrementa l’ampiezza delle onde lente favorendo la generazione di potenziali d’azione e la forza di contrazione La stimolazione simpatica ha un effetto inibitorio sulla muscolatura liscia, NB una azione eccitatoria si manifesta solo a livello della muscolaris mucosae e degli sfinteri