Photosynthesis Photosynthesis is the process of converting energy



























 were created in step")






 accumulate inside thylakoids. • The light reactions of photosynthesis occur")















.")


- Slides: 57
Photosynthesis
• Photosynthesis is the process of converting energy from sunlight to energy in chemical bonds. • The general equation describing photosynthesis is Carbon dioxide Water Glucose Oxygen gas • C 6 H 12 O 6 is glucose. • Sometimes you will see CH 2 O or (CH 2 O)n. These are general formulas for glucose or any carbohydrate.
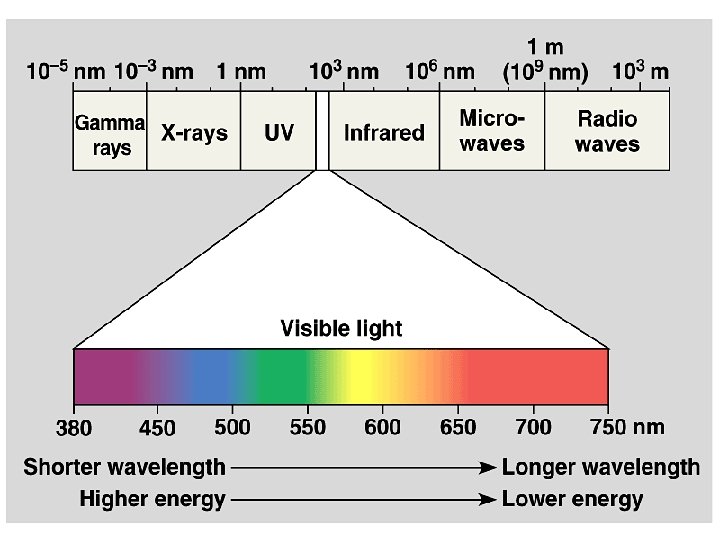
• The process of photosynthesis begins with light-absorbing pigments in plant cells. • A pigment molecule is able to absorb the energy from light only within a narrow range of wavelengths. • In order to absorb as much of the entire bandwidth from sunlight as possible, different pigments, capable of absorbing different wavelengths, act together to optimize energy absorption. • These pigments include the green chlorophyll a and chlorophyll b and the carotenoids, which are red, orange, or yellow.
• When the light is absorbed into one of these pigments, the energy from the light is incorporated into electrons within the atoms that make up the molecule. • These energized electrons (or “excited” electrons) are unstable and almost immediately re-emit the absorbed energy. • The energy is then reabsorbed by electrons of a nearby pigment molecule. Energy reabsorbed by electrons of a nearby pigment molecule
• The process of energy absorption, followed by re-emission of energy, continues, with the energy bouncing from one pigment molecule to another.
• The process ends when the energy is absorbed by one of two special chlorophyll a molecules, P 680 and P 700.
• These two chlorophyll molecules, named with numbers that represent the wavelengths at which they absorb their maximum amounts of light (680 and 700 nanometers), are different from other chlorophyll molecules because of their association with various nearby pigments.
• Together with these other pigments, chlorophyll P 700 forms a pigment cluster called photosystem I (PS I). • Chlorophyll P 680 forms photosystem II (PS II). Primary electron acceptor Photon “Reaction center” chlorophyll P 700 or P 680 Pigment molecules PHOTOSYSTEM
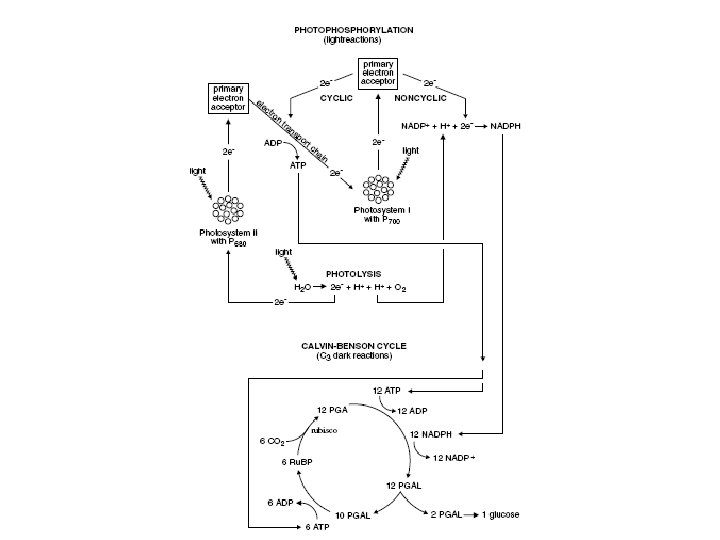
• The chemical reactions that now take place are illustrated in the Figure. • Refer to the figure as you read the following descriptions of the metabolic processes.
• During the reactions, there are two possible routes for electron flow: • cyclic • and noncyclic.
Noncyclic Photophosphorylation • Photophosphorylation is the process of making ATP from ADP and Pi (phosphorylation) using energy derived from light (photo). • Noncyclic photophosphorylation begins with PS II and follows the steps: Calvin-Benson Cycle
Noncyclic Photophosphorylation • 1. Photosystem II. Electrons trapped by P 680 in photosystem II are energized by light. • The two electrons are “moving” up, signifying an increase in their energy. Calvin-Benson Cycle
Noncyclic Photophosphorylation • 2. Primary electron acceptor. • Two energized electrons are passed to a molecule called the primary electron acceptor. • This electron acceptor is called “primary” because it is the first in a chain of electron acceptors. Calvin-Benson Cycle
Noncyclic Photophosphorylation • 3. Electron transport chain. • Electrons pass through an electron transport chain. • This chain consists of proteins that pass electrons from one carrier protein to the next. • Some carrier proteins, like ferredoxin and cytochrome, include nonprotein parts containing iron. Calvin-Benson Cycle
Noncyclic Photophosphorylation • 4. Phosphorylation. • As the two electrons move “down” the electron transport chain, they lose energy. • The energy lost by the electrons as they pass along the electron transport chain is used to phosphorylate, on average, about 1. 5 ATP molecules. Calvin-Benson Cycle
Noncyclic Photophosphorylation • 5. Photosystem I. • The electron transport chain terminates with PS I (with P 700). • Here the electrons are again energized by sunlight and passed to a primary electron acceptor (different from the one associated with PS II). Calvin-Benson Cycle
Noncyclic Photophosphorylation • 6. NADPH. • The two electrons pass through a short electron transport chain. • At the end of the chain, the two electrons combine with NADP+ and H+ to form NADPH. • NADPH is a coenzyme. Since the electrons have a considerable amount of energy left, NADPH is an energy-rich molecule. Calvin-Benson Cycle
Noncyclic Photophosphorylation • 7. Photolysis. • The two electrons that originated in PS II are now incorporated into NADPH. • The loss of these two electrons from PS II is replaced when H 2 O is split into two electrons, 2 H+ and 1⁄2 O 2. • The process is called photolysis and literally means decomposition (lysis) by light (photo). • A manganese-containing protein complex catalyzes the reaction. • The two electrons from H 2 O replace the lost electrons from PS II. • One of the H+ provides the H in NADPH. Calvin-Benson Cycle
• In summary, photophosphorylation takes the energy in light and the electrons in H 2 O to make the energy-rich molecules ATP and NADPH. • Because the reactions require light, they are often called the light-dependent reactions or, simply, light reactions. • The following equation informally summarizes the process: H 2 O + ADP + Pi + NADP+ + light ATP + NADPH + O 2 + H+
El Primary electron acceptor ec tro n tra ns po rt El ec tro n tra ns po r tc ha in Photons Energy for synthesis of PHOTOSYSTEM II by chemiosmosis
Cyclic Photophosphorylation • A second photophosphorylation sequence occurs when the electrons energized in PS I are “recycled. ” • In this sequence, energized electrons from PS I join with protein carriers and generate ATP as they pass along the electron transport chain. Calvin-Benson Cycle
Cyclic Photophosphorylation • In contrast to noncyclic photophosphorylation where electrons become incorporated into NADPH, electrons in cyclic photophosphorylation return to PS I. • Here they can be energized again to participate in cyclic or noncyclic photophosphorylation. • Cyclic photophosphorylation is considered a primitive form of photosynthesis but occurs simultaneously with noncyclic photophosphorylation. Calvin-Benson Cycle
PHOTOSYSTEM I with P 700
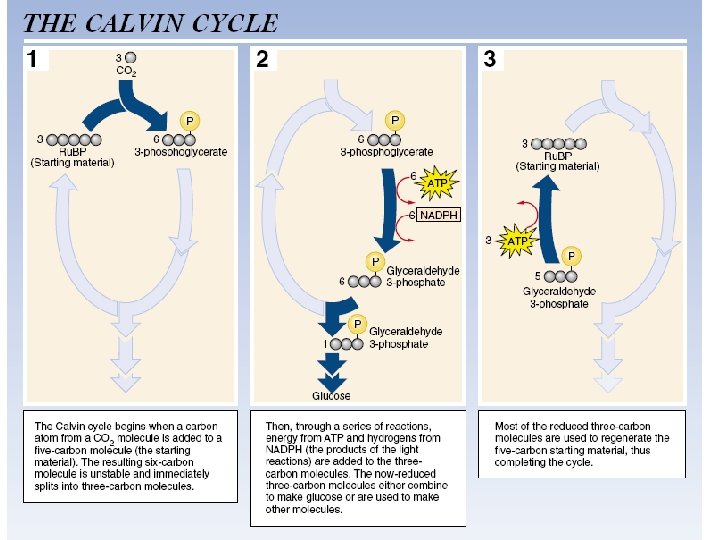
Calvin-Benson Cycle • The Calvin-Benson cycle “fixes” CO 2. That is, it takes chemically unreactive, inorganic CO 2 and incorporates it into an organic molecule that can be used in biological systems. • The biosynthetic pathway involves over a dozen products. • The function of the pathway is to produce a single molecule of glucose (C 6 H 12 O 6). • In order to accomplish this, the Calvin-Benson cycle must repeat six times, and use 6 CO 2 molecules. • Thus, in the Figure and the discussion that follows, all the molecules involved have been multiplied by 6. • Only the most important molecules are discussed.
1. Carboxylation: 6 CO 2 combine with 6 Ru. BP to produce 12 PGA. • The enzyme Ru. BP carboxylase, or rubisco, catalyzes the merging of CO 2 and Ru. BP (ribulose bisphosphate). • The Calvin-Benson cycle is referred to as C 3 photosynthesis because the first product formed, PGA (phosphoglycerate), contains three carbon atoms. • Other names are the Calvin cycle and the carbon reduction cycle.
2. Reduction: 12 ATP and 12 NADPH are used to convert 12 PGA to 12 PGAL. • The energy in the ATP and NADPH molecules is incorporated into PGAL (glyceride 3 -phosphate), thus making PGAL a very energyrich molecule. • ADP, Pi, and NADP+ are released and then re-energized in noncyclic photophosphorylation.
3. Regeneration: 6 ATP are used to convert 10 PGAL to 6 Ru. BP. Regenerating the 6 Ru. BP originally used to combine with 6 CO 2 allows the cycle to repeat.
4. Carbohydrate synthesis. Note that 12 PGAL (glyceride 3 -phosphate) were created in step 2, but only 10 were used in step 3. What happened to the remaining two? These two remaining PGAL are used to build glucose, a common energystoring molecule. • Other monosaccharides like fructose and maltose can also be formed. • In addition, glucose molecules can be combined to form disaccharides like sucrose and polysaccharides like starch and cellulose.
• Details of the Calvin cycle 6 INPUT: Step 1 Carboxylation CO 2 In a reaction catalyzed by rubisco, 6 molecules of CO 2 are fixed. 1 6 P P 12 P Ru. BP Step 2 Reduction. 3 -PGA 12 ATP 6 ADP 6 ATP CALVIN CYCLE 3 Step 3 Regeneration of Ru. BP. 12 ADP + P 2 12 NADPH 12 NADP+ 10 P 12 P G 3 P Step 4 Carbohydrate synthesis. 4 OUTPUT: 2 P G 3 P Glucose and other compounds
• You should recognize that no light is directly used in the Calvin-Benson cycle. Thus, these reactions are often called the light-independent reactions or even the dark reactions. • But be careful—the process cannot occur in the dark. This is because it is dependent upon the energy from ATP and NADPH, and these two energy-rich molecules can be created only during photophosphorylation, which can occur only in light.
• In summary, the Calvin-Benson cycle takes CO 2 from the atmosphere and the energy in ATP and NADPH to create a glucose molecule. • Of course, the energy in ATP and NADPH represents energy from the sun captured during photophosphorylation. • The Calvin-Benson cycle can be informally summarized as follows: INPUT 6 CO 2+18 ATP+12 NADPH + H+ 18 ADP + 18 Pi + +12 NADP + 1 glucose • Keep in mind that the reactions that occur during photosynthesis (and in any biosynthetic pathway) do not occur spontaneously. • Every product formed in every reaction is catalyzed by an enzyme. • In some reactions, coenzymes or metal-ion cofactors may also be involved. CALVIN CYCLE OUTPUT:
Chloroplasts • Chloroplasts are the sites where both the light-dependent and lightindependent reactions of photosynthesis occur. • Chloroplasts consist of an outside phospholipid bilayer membrane enclosing a fluid called the stroma. • Suspended within the stroma are stacks of pancakelike membranes. Individual membrane layers (the “pancakes”) are thylakoids; an entire stack of thylakoids is a granum (plural, grana). • Within the thylakoids are the light-absorbing pigments and enzymes for the light-dependent reactions. • In the surrounding stroma are the enzymes for the Calvin-Benson cycle. • Thus, the light reactions occur on the thylakoid membranes, and the dark reactions occur in the stroma.
• The location and structure of chloroplasts Chloroplast LEAF CROSS SECTION MESOPHYLL CELL LEAF Mesophyll CHLOROPLAST Intermembrane space Outer membrane Granum Grana Stroma Inner membrane Stroma Thylakoid compartment
Chemiosmotic Theory • Chemiosmotic theory describes the mechanism by which ADP is phosphorylated to ATP. • The process illustrated in the Figure follows the steps listed below.
1. H+ ions (protons) accumulate inside thylakoids. • The light reactions of photosynthesis occur in the membranes of the thylakoids. • As photolysis occurs, H+ are created and released to the inside of the thylakoids (the oxygen is released to the outside). • Also, H+ accompany the electrons as they pass along the electron transport chain between PS II and PS I. • These H+ come from the stroma (outside thylakoids) and are released to the inside of the thylakoids.
2. A p. H and electrical gradient across the thylakoid membrane is created. • As H+ accumulate inside thylakoid, the p. H decreases. • Since some of these H+ come from outside thylakoids (from the stroma), the H+ concentration decreases in the stroma and its p. H increases. • This creates a p. H gradient consisting of differences in the concentration of H+ across the thylakoid membrane from a stroma p. H 8 to a thylakoid p. H 5. • Since H+ are positively charged, their accumulation on the inside of the thylakoid creates an electric gradient (or voltage) as well.
3. ATP synthases generate ATP. • The p. H and electrical gradient represent potential energy like water behind a dam. • Similar to a dam, channel proteins, called ATP synthases, allow the H+ to flow through the thylakoid membrane and out to the stroma. • The energy generated by the passage of the H+ (like the water through turbines in a dam) provides the energy for the ATP synthases to phosphorylate ADP to ATP. • The passage of about three H+ is required to generate one ATP.
Fig. 10. 16 Copyright © 2002 Pearson Education, Inc. , publishing as Benjamin Cummings
Photorespiration • Because of its critical function in catalyzing the fixation of CO 2 in all photosynthesizing plants, rubisco is the most common protein on earth. However, it is not a particularly efficient molecule. • In addition to its CO 2 -fixing capabilities, it is also able to fix oxygen. • The biosynthetic pathway that leads to the fixation of oxygen is called photorespiration. CALVIN CYCLE 2 -C compound peroxisomes
Photorespiration leads to two problems. – The first is that the CO 2 -fixing efficiency is reduced because, instead of fixing only CO 2, rubisco fixes some O 2 as well. – The second problem is that the products formed when O 2 is combined with Ru. BP (ribulose bisphosphate) do not lead to the production of useful, energy-rich molecules like glucose. Instead, specialized cellular organelles, the peroxisomes, are found near chloroplasts, where they function to break down photorespiration products. Thus, considerable effort is made by plants to rid the cell of the products of photorespiration. Since the early atmosphere in which primitive plants originated contained very little oxygen, it is hypothesized that the early evolution of rubisco was not influenced by its O 2 -fixing handicap ( )ﻋﺎﺋﻖ.
Photorespiration usually occurs when oxygen levels are high; for example, when the stomata are closed to prevent water loss on dry days. To prevent this process, two specialized biochemical mechanisms have been evolved in the plant world to reduce uptake of molecular oxygen by Rubisco: C 4 and CAM metabolism.
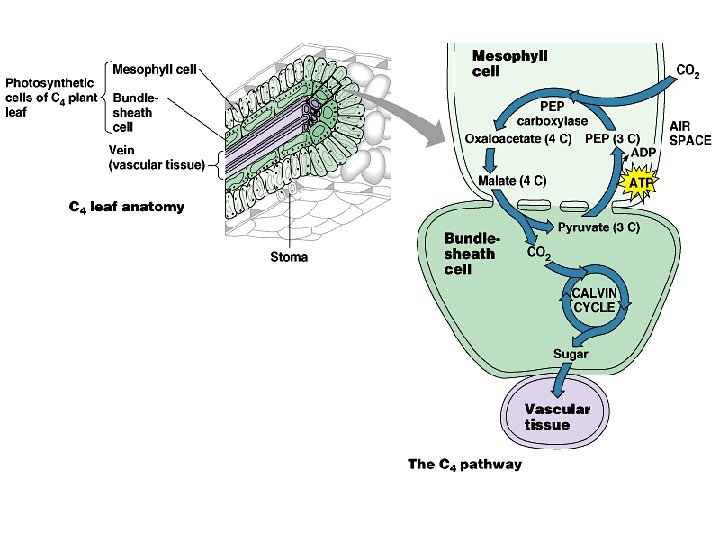
C 4 Photosynthesis • Improving upon photosynthetic efficiency, some plants have evolved a special “add-on” feature to C 3 photosynthesis. • When CO 2 enters the leaf, it is absorbed by the usual photosynthesizing cells, the mesophyll cells. • Instead of being fixed by rubisco into PGA (phosphoglycerate), the CO 2 combines with PEP (phosphoenolpyruvate) to form OAA (oxaloacetate or oxaloacetic acid). • The fixing enzyme is PEP carboxylase.
• OAA, the first product of this pathway, has 4 carbon atoms, thus the name C 4 photosynthesis. • OAA is then converted to malate. oxaloacetate
• The malate is shuttled to specialized cells within the leaf, the bundle sheath cells. • Here malate is converted to pyruvate and CO 2. • The pyruvate is then shuttled back to the mesophyll cells where one ATP (broken down to AMP, instead of ADP) is required to convert the pyruvate back to PEP. • Then the process repeats. • The overall effect is to move CO 2 from mesophyll cells to the bundle sheath cells. oxaloacetate
• The purpose for moving CO 2 to bundle sheath cells is to increase the efficiency of photosynthesis. • The bundle sheath cells surround the leaf veins and are themselves surrounded by densely packed mesophyll cells. • Since bundle sheath cells rarely make contact with an intercellular space, very little oxygen reaches them.
• When malate delivers CO 2 to a bundle sheath cell, rubisco begins the Calvin-Benson cycle (C 3 photosynthesis). • Because little oxygen is present, rubisco can fix CO 2 without competition from O 2. • Thus, little photorespiration takes place, and photosynthesis is more efficient.
• In order for photosynthesis to occur, the stomata must be open to allow CO 2 to enter. • However, when the stomata are open, H 2 O can escape. • The higher rate of photosynthesis among C 4 plants allows them to reduce the time that the stomata are open, thereby, reducing H 2 O loss.
• Thus, C 4 plants are found in hot, dry climates, where they possess an adaptive advantage over C 3 plants. • This advantage apparently compensates for the additional energy requirement (1 ATP to AMP) for C 4 photosynthesis. • C 4 photosynthesis occurs in about a dozen plant families. Sugarcane and crab grass are two examples. crab grass Sugarcane
CAM Photosynthesis • Another “add-on” feature to C 3 photosynthesis is crassulacean acid metabolism (CAM) • The physiology of this pathway is almost identical to C 4 photosynthesis, with the changes that follow:
1. PEP carboxylase still fixes CO 2 to OAA, as in C 4. Instead of malate, however, OAA is converted to malic acid. (This is a minor difference, since malate is merely the ionized form of malic acid). 2. Malic acid is shuttled to the vacuole of the cell (not moved out of the cell to bundle sheath cells as in regular C 4). 3. Stomata are open at night. During the night, PEP carboxylase is active and malic acid accumulates in the cell’s vacuole.
• 4. Stomata are closed during the day (the reverse of other plants). • At this time, malic acid is shuttled out of the vacuole and converted back to OAA (requiring 1 ATP to ADP), releasing CO 2. • The CO 2 is now fixed by rubisco, and the Calvin-Benson cycle proceeds.
• The advantage of CAM is that photosynthesis can proceed during the day while the stomata are closed, greatly reducing H 2 O loss. • As a result, CAM provides an adaptation for plants that grow in hot, dry environments with cool nights (such as deserts).
• The name crassulacean acid metabolism comes from the early discovery of CAM in the succulent plants of the family Crassulaceae and the discovery of the accumulation of malic acid in vacuoles during the night. • In addition to the Crassulaceae, CAM is found among plants in over a dozen different families, including cacti. Crassula ovata