Cours 2 ANATOMIE DE LA SENSATION SOMATIQUE INTRODUCTION







 La matière blanche contient des axones myélinisés La matière blanche est divisée en")
 Les fibres des racines dorsales parcourent la matière blanche et se diffusent dans")

 et")
 Le")


 Le système antéro-latéral contrôle la perception de la douleur et de la température")







- Slides: 28
Cours 2 ANATOMIE DE LA SENSATION SOMATIQUE
INTRODUCTION Différents récepteurs sensoriels somatiques sont sensibles à différentes qualités de stimuli. Cette spécificité est partagée par les chaînes de neurones qui transportent l’information sensorielle par le biais de faisceaux ascendants vers le cortex cérébral. Les fibres afférentes se connectent sur des régions bien particulières aussi bien au niveau de la ME que du TC. A partir de là, l’information remonte jusqu’au thalamus, avant d’être transmise jusqu’au cortex cérébral.
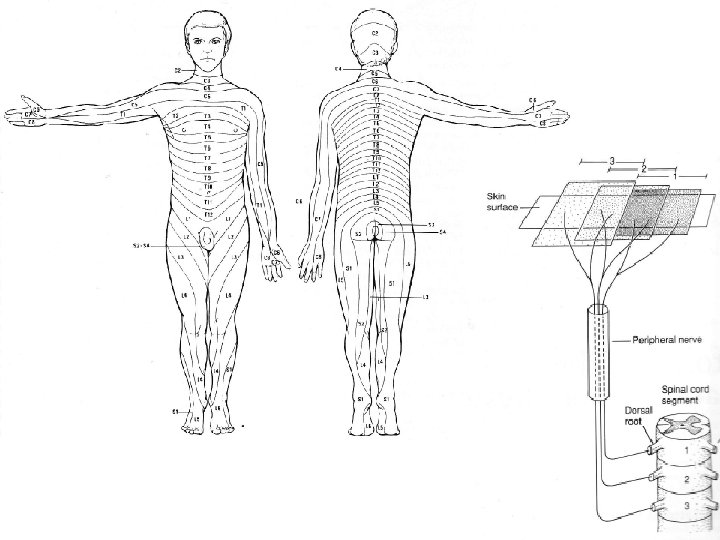
I LE TERRITOIRE CUTANE INNERVE PAR UNE RACINE DORSALE EST APPELE DERMATOME L’information périphérique est amenée au SNC par des fibres afférentes qui sont mélangées avec les fibres efférentes dans les nerfs périphériques. En se rapprochant de la ME, on assiste à une séparation qui permet aux fibres afférentes de pénétrer au niveau des racines dorsales. Ainsi, chaque racine dorsale innerve une région de la périphérie bien particulière : le dermatome. Toutefois, du fait de convergences au niveau des plexus, les territoires couverts par les différentes et successives racines dorsales se recouvrent largement.
Toutefois, la section d’une portion distale d’un nerf cutané périphérique a pour effet la perte de sensation cutanée sur une partie limitée de la peau. En revanche, un nerf spinal ou une racine dorsale endommagée n’occasionne souvent qu’un déficit sensoriel mineur. Puisque les fibres d’une racine dorsale transmettent plusieurs modalités sensorielles (tactile, proprioception, douleur, température), les limites dermatomes peuvent être explorées en recourant à divers stimuli. Les tests utilisés pour localiser les limites de ces territoires permettent de mettre en évidence une variabilité en fonction de la nature du stimulus.
Une autre méthode pour étudier les zones distribuées par une racine dorsale et de regarder le territoire lésionnel résultant d’un zona, une inflammation douloureuse des ganglions des racines dorsales par une infection virale et s’accompagnant d’une éruption limitée au dermatome correspondant.
II LA MOELLE EPINIERE EST COMPOSEE DE MATIERES BLANCHE ET GRISE Pour transporter l’information jusqu’au cerveau (-> perception), les voies sensorielles somatiques doivent au préalable traverser la ME. Cette dernière a en fait 3 fonctions identifiables : - servir de relais pour transmettre l’information sensorielle - amener les voies ascendantes afférentes et les voies motrices descendantes commandant le tronc et les jambes - contenir les Ins et Mns qui contrôlent la motricité du tronc et des jambes
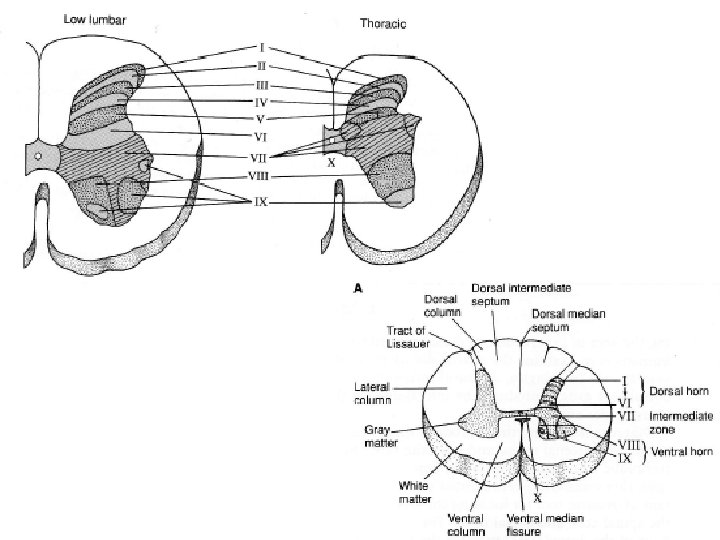
Sur une section schématique, on peut constater, outre une symétrie évidente, que la ME est composée de matière grise, qui contient les corps cellulaires des neurones, et de matière blanche, qui contient les faisceaux. 1) La matière grise spinale contient des corps cellulaires de nerfs La forme en aile de papillon qui caractérise la matière grise est divisée en une corne dorsale, une zone intermédiaire et une corne ventrale. Ces zones peut être subdivisée en noyaux ou couches. - les I à V sont correspondent à la corne dorsale, - les VI et VII à la zone intermédiaire et VIII et IX à la corne ventrale. - la X à la matière grise entourant le canal central.
2) La matière blanche contient des axones myélinisés La matière blanche est divisée en 3 paires de colonnes : - les colonnes dorsales qui relaient les informations sensorielles somatiques vers le bulbe rachidien (BR). - les colonnes latérales qui contiennent les axones descendant du cerveau qui contrôlent des fonctions aussi bien sensorielles que motrices ainsi que des faisceaux sensoriels ascendants. - les colonnes ventrales qui contiennent les neurones moteurs des muscles squelettiques. En supplément de ces colonnes, on trouve des neurones propriospinaux qui assurent les connexions entre diverses régions de la ME.
3) Les fibres des racines dorsales parcourent la matière blanche et se diffusent dans la matière grise Parmi les fibres qui entrent dans la ME, les plus grandes, les myélinisées, peuvent avoir jusqu’à 20 microns de diamètre et pénètrent de façon médiane (entre les cornes dorsales). Les plus petites, non myélinisées, peuvent avoir moins de 1 micron de diamètre et pénètrent dans la ME de façon plus latérale. Une fois dans la ME, les fibres remontent et descendent dans la matière blanche pour de distribuer dans la matière grise. Selon les modalités sensorielles, les axones ont des distributions différentes.
Les fibres de petits diamètres, qui transmettent les sensations de douleur et de température, n’entrent pas immédiatement dans la matière grise mais transitent par la couche I. Là les fibres bifurquent vers le haut ou vers le bas pour se terminer au niveau des couches I, II et III. Les fibres de plus gros diamètres, qui transmettent le sens tactile et la proprioception segmentaire entrent d’une façon médiane entre les cornes dorsales.
Certaines fibres de gros diamètres se terminent sur des noyaux moteurs (couche IX) et contrôlent le réflexe d’étirement (réflexe intrasegmentaire : les fibres afférentes collatérales entrent et se terminent à un même niveau de la ME). A l’inverse, les réflexes de grattage et les réflexes posturaux consécutifs à une perturbation constituent des exemples de réflexes intersegmentaux. Les neurones qui véhiculent le sens du toucher se terminent sur le noyau propre (couches III et IV) alors que les afférences proprioceptives se terminent plus ventralement (couche VII), sur les Ins des couches V et VI et sur les Mns de la couche IX.
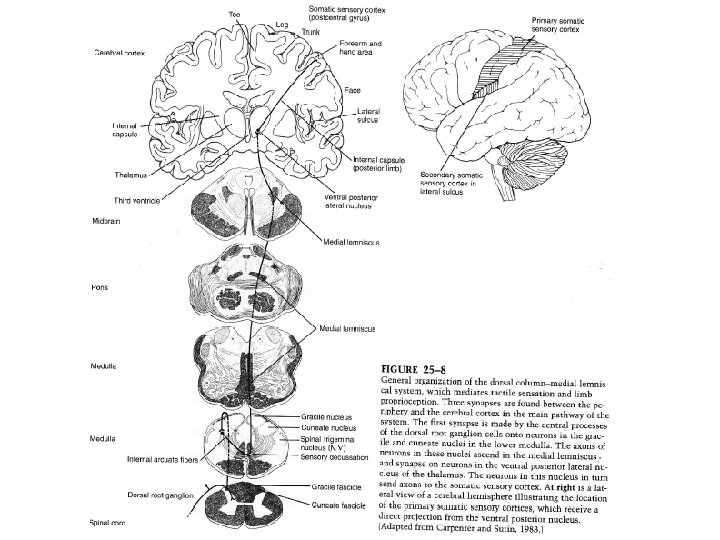
III DEUX SYSTEMES ASCENDANTS MAJEURS TRANSPORTENT L’INFORMATION SENSORIELLE VERS LE CORTEX CEREBRAL 1) Le système colonne dorsale- lemnisque médian assure le sens tactile et la proprioception segmentaire Les colonnes dorsales sont composées principalement (85%) des branches périphériques qui remontent jusqu’à la base du TC sans faire synapse. 15% des fibres des colonnes dorsales sont des axones ascendants des neurones des cornes dorsales. Aux plus hauts niveaux de la ME, ces colonnes dorsales peuvent être divisées en deux faisceaux gracile (Goll) et cunéiforme (Burdach).
Les afférences des segments ipsilatéraux sacrés, lombaires et thoraciques bas remontent selon une position médiane dans le faisceau gracile. Les afférences des segments thoraciques hauts et cervicaux remontent plus latéralement dans le faisceau cunéiforme. Les fibres quittent les noyaux des colonnes dorsales ont une trajectoire arrondie lorsqu’elles traversent la ligne médiane, d’où leur nom de fibres arquées internes ou faisceau en virgule. Après décussation, elles rejoignent le lemnisque médian et remontent jusqu’au thalamus Bien que les colonnes dorsales contiennent à la fois des axones tactiles et proprioceptifs, ces deux sousmodalités demeurent anatomiquement séparées.
Les axones des propriocepteurs sont positionnés plus ventralement dans les colonnes dorsales que ceux des récepteurs tactiles qui sont situés dorsalement. Au niveau du BR, les fibres décussent et appartiennent au lemnisque médian. Au niveau du thalamus, les fibres lemniscales font synapse avec les neurones du VPL. Les afférences sensorielles de la face font synapse avec le VPM.
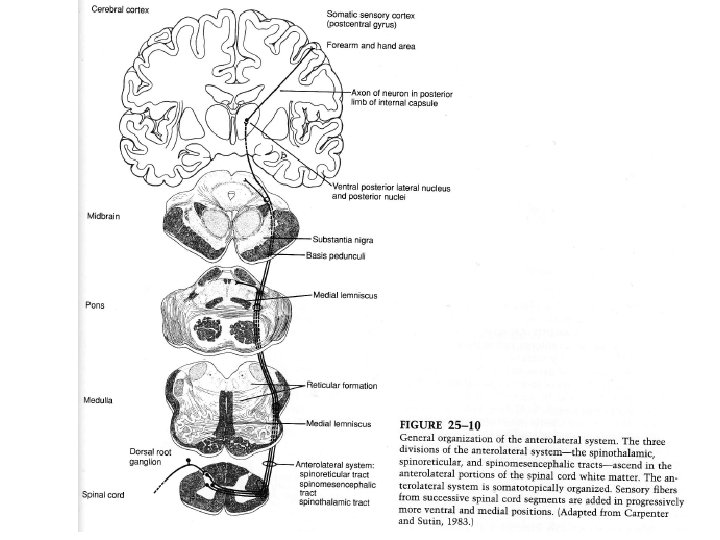
2) Le système antéro-latéral contrôle la perception de la douleur et de la température Il est composé de voies séparées qui jouent un rôle majeur dans la perception de la douleur et de la température et un rôle mineur dans le sens tactile et la proprioception. Ce système diffère du précédent système (lemniscal médian et colonnes dorsales) pour plusieurs raisons : - Le système antéro-latéral (SAL) possède un important contingent de fibres ascendantes qui ne décussent pas - Lorsqu’elle intervient, la décussation s’effectue au niveau de la ME (BR pour le lemnisque médian)
- Les cellules d’origine du SAL se situent dans la corne dorsale et de ce fait sont des neurones post-synaptiques vis à vis des fibres afférentes primaires. A l’inverse, les axones des colonnes dorsales sont le plus souvent des collatérales des fibres afférentes primaires. La plupart des fibres du lemnisque médian se terminent sur le thalamus alors que celle du SAL se terminent sur le thalamus et le TC. Le SAL est composé de trois faisceaux principaux qui se distinguent par leur site de terminaison : Le faisceau spino-thalamique a son origine principalement au niveau de la couche I de la ME, mais aussi des couches V à VII. Il assure le transport rapide de la douleur et de température par des fibres Ad jusqu’au VPL thalamique.
- le faisceau spino-réticulaire est important pour la douleur lente, avec une conduction assurée par des fibres C. Ce faisceau, qui naît au niveau des couches VII et VIII se termine sur des neurones de la formation réticulée du TC, qui ensuite relaient cette information à des niveaux plus hauts (thalamus et autres structures du diencéphale comme l’hypothalamus). - Le faisceau spino-mésencéphalique se termine au niveau du tectum (colliculi) et de la matière grise périaqueducale. Les fibres du SAL font synapse sur des noyaux de trois régions thalamiques : les noyaux postérieurs, les noyaux intra-laminaires et les noyaux VPL.
Les VPL se projettent sur les aires corticales sensorielles somatiques. Les noyaux intra-laminaires se projettent plus largement sur le TC et le cortex. Les noyaux postérieurs se projettent sur des régions du lobe pariétal en dehors des aires sensorielles somatiques. Conséquences fonctionnelles de cette organisation: Après une hémisection médullaire thoracique, le sens tactile et la proprioception, relayés par les colonnes dorsales, sont perdus sur le côté ipsilatéral alors que la douleur et le sens thermique, relayés par le SAL, sont perdus du côté controlatéral.
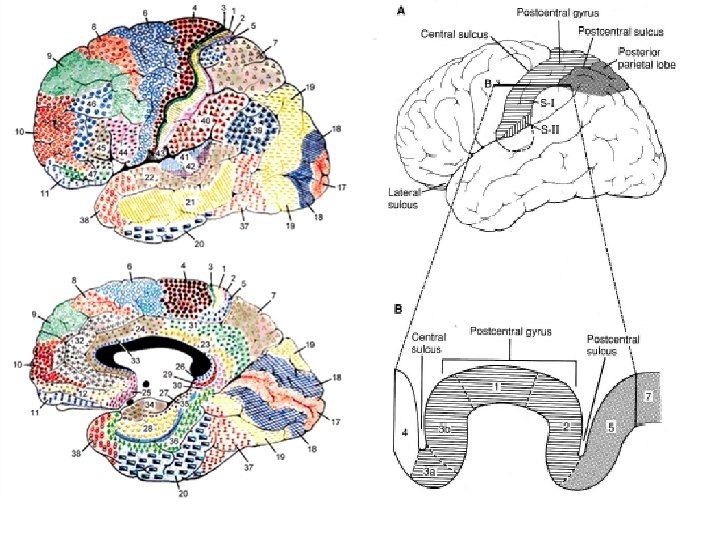
IV LE CORTEX SENSORIEL SOMATIQUE PRIMAIRE EST DIVISE EN QUATRE PARTIES Plusieurs régions distinctes du cortex pariétal antérieur reçoivent des informations somatiques. L’information relayée vers ces aires corticales est surtout importante pour le sens tactile et la proprioception. Le cortex sensoriel somatique primaire (S-I) est situé dans la circonvolution post-centrale et dans les profondeurs de la scissure centrale de Rolando. Il est constitué des aires de Brodmann (AB) 1, 2, 3 a et 3 b. Située en arrière et plus latéralement de S-I se trouve le cortex sensoriel somatique secondaire (S-II).
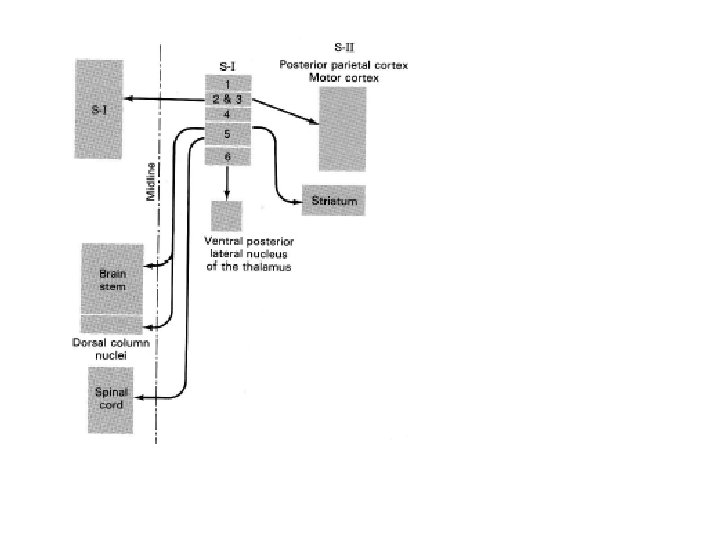
Chacune de ces deux aires est organisée somatotopiquement. Les signaux afférents allant vers S-I proviennent de la partie du corps controlatérale, alors que ceux qui se dirigent vers S-II sont bilatéraux. Les projections thalamiques directes vers S-I et S-II proviennent essentiellement du noyau ventro-postérieur (VPL et VPM). Une troisième région corticale (lobe pariétal postérieur; AB 5 et 7) reçoit des signaux afférents somatiques. Cette région est assimilée à une aire associative en ce sens qu’elle relie des processus moteurs et sensoriels et qu’elle est impliquée dans l’intégration de différentes modalités sensorielles nécessaires à la perception.
V LES CELLULES PYRAMIDALES CONSTITUENT LA SORTIE DU CORTEX CEREBRAL Les neurones du cortex sensoriel somatique qui reçoivent des signaux thalamiques font quatre types de connections : intra-corticales, associatives, calleuses et sub-corticales. Les connections intra-corticales sont faites vers des neurones du cortex : les cellules des grains qui, avec les cellules pyramidales, constituent les deux principaux types de neurones corticaux. La densité de ces deux types de cellule dans chacune des couches corticales est inégale. Les couches riches en cellules pyramidales sont efférentes alors que celles riches en cellules à grains voient converger les afférences (essentiellement thalamiques).
Les connections d’association permettent de communiquer entre différentes régions corticales du même côté. Il existe de telles connections entre chacune des AB 1, 2 et 3 (S-I), entre S-I et S-II, et entre les cortex sensoriels primaires et secondaires et l’AB motrice n° 4. Les connections calleuses mettent en relation des aires symétriques deux hémisphères. Quatre importantes projections descendantes de S-I sont réalisées : vers les ganglions de la base (striatum), vers le noyau thalamique VP, vers les noyaux des colonnes dorsales et vers les cornes dorsales de la ME. Ces projections permettent un contrôle cortical des messages sensoriels, ce qui constitue un élément important de l’organisation sensorielle.