KARYOTK HCRE DNGSNN DZENLENMES Prof Dr Asuman Sungurolu






 Restriksiyon noktası yer alır. Hücrede bölünmeyi uyaran büyüme")



 ORGANİZMA / PROTEİN ADI S.")















 ailesi ve INK")
- Slides: 28
ÖKARYOTİK HÜCRE DÖNGÜSÜNÜN DÜZENLENMESİ Prof. Dr. Asuman Sunguroğlu
Hücreler neden bölünür? n Büyüme n Tamir/Yenilenme n Üreme – Aseksüel
Hücrenin yaşam döngüsü Hücre bölünmesi – Mitoz – 1 saat İkinci büyüme fazı – 4 saat interphase Sentez fazı – DNA kopyalanır – 8 saat Birinci büyüme fazı, 11 saat
HÜCRE DÖNGÜSÜ G 1 = Yeni hücre meydana geldikten sonra hücre büyür. n S – DNA replikasyonu gerçekleşir. n G 2 – Mitoz bölünme için hazırlık evresidir, hücre büyümeye devam eder. n M – Mitoz: Birbirinin aynısı 2 hücre meydana gelir. n Eğer hücre bölünmeyecekse G 0 fazı olarak adlandırılan dinlenme fazında bekler – Hücre bölünmesi için hazırlık yapılmaz. n
Hücrede G 1 in sonlarında G 2’nin sonlarında ve Mitozda kontrol noktaları bulunur.
Mayada G 1’in sonlarındaki kritik bir nokta vardır, bu START noktasını geçince hücre bölünmesi mitoz tamamlanıncaya kadar devam eder.
Mayalardaki START noktasının eşdeğeri olarak memelide(insanda) Restriksiyon noktası yer alır. Hücrede bölünmeyi uyaran büyüme faktörleri veya mitojen varlığında hücre G 0 dan G 1’e geçer ve bölünmeye başlar.
Büyüme faktörleri uayrımını alan hücrede RAS/RAF/ERK Kinaz yolağı aktive olur ve ilk olarak Siklin D proteini sentezlenir.
n Yaban-tip S. pombe genleri, üzerinde artı işaretiyle italik olarak yazılır (ör; cdc 2 +); çekinik mutasyonlu genler ise üzerinde eksi işaretiyle italik olarak yazılır (ör; cdc 2 -). Belirli genler tarafından kodlanan proteinler, genin sembolü ile isimlendirilir ve ilk harfi büyük olacak şekilde roman harfleriyle yazılarak gösterilir (ör; Cdc 2).
n S. cerevisiae yaban tip genler, S. pombe’de olduğu gibi, italik büyük harflerle gösterilirler (ör; CDC 28) ve çekinik mutant genler, italik küçük harflerle gösterilirler (ör; cdc 28); yaban tip proteinler Romen harfleriyle ilk harfi büyük olarak yazılır (ör; Cdc 28).
TABLO 20 -1 Seçilmiş Siklinler ve Siklin-Bağımlı Kinazlar (CDK’lar) ORGANİZMA / PROTEİN ADI S. POMBE CDK Cdc 2 Mitotik Siklin Cdc 13 S. CEREVISIAE CDK Cdc 28 Orta-G 1 siklin Cln 3 Geç-G 1 siklinleri Cln 1, Cln 2 Erken S-faz siklinleri Clb 5, Clb 6 Geç S-faz ve erken mitotik siklinler Clb 3, Clb 4 Geç mitotik siklinler Clb 1, Clb 2 OMURGALILAR Orta-G 1 CDK 4, CDK 6 Geç-G 1 ve S-faz CDK 2 Mitotik CDK’lar CDK 1, CDK 2 Orta-G 1 siklinler D-tipi siklinler Geç-G 1 ve S-faz siklin Siklin E S-faz ve mitotik siklin Siklin A Mitotik siklinler Siklin A, Siklin B
Moleküler Hücre Biyolojisi Lodish 2011
Moleküler Hücre Biyolojisi Lodish 2011
Aktif MPF P Baskılayıcı fosforilasyon Moleküler Hücre Biyolojisi Lodish 2011 Aktifleştiren fosforilasyon
MPF: Maturation promoting factor Mitotik Siklin olarak sentezlenen Siklin. B Siklin bağımlı Kinaz ile kompleks yapar ve Kinaz aktivasyonu gerçekleşir. Kinazın aktivasyonu için Thr 161, baskılanması için Thr 14 ve Tyr 15 e fosfat takılması gereklidir.
MPF MPF, aktivitesi için mitotik sikline ihtiyaç duyan bir protein kinazdır. MPF’nin protein kinaz aktivitesi, birçoğu hala tanımlanmayı bekleyen birçok spesifik protein substratın fosforlanmasıyla mitozun başlamasını uyarır.
Molecular Cell Biology Alberts, 3 rd edition
Kondensin komplekslerinin MPF veya MPF tarafından düzenlenen bir kinaz tarafından fosforilasyonu erken mitozda kromozom yoğunlaşmasını sağlar.
n Mitozun başlarında, lamin A, B ve C’nin, nükleoporinlerin ve iç nükleus zar proteinlerinin MPF-tarafından fosforilasyonu, lamin filamentlerinin depolimerizasyonuna, nükleer zarfın bütünlüğünü yitirmesine ve ER’a çekilmesine yol açan nükleer porların por alt komplekslerine ayrılmasına sebep olur.
Sccafold maintenance kompleks Moleküler Hücre Biyolojisi Lodish 2011
Telofaz esnasında nükleer zarın yeniden toplanması için model. Her bir seyrelen ve daha sonra biri diğerine füzyon yapan kromozomla ilişkideki endoplazmik retikulumun (ER) uzantıları kromozomun etrafında çift zar oluşturur. Fosfatı kopan nükleer por alt kompleksleri nükleer porlarda yeniden toplanarak karyomer olarak adlandırılan bireysel mininükleus oluştururlar. Çevrili kromozom daha fazla seyrelir ve her bir iğ kutbundaki tüm karyomerlerin nükleer zarlarının takip eden füzyonu, tam set kromozom içeren tek nükleusu oluşturur. NPC = nükleer por kompleks. [B. Burke ve J. Ellenberg, 2002, Nature Rev. Mol. Cell Biol. 3: 487’den adapte edilmiştir. ]
n S-faz siklin-CDK heterodimerleri geç G 1’de birikirken, geç mitozda ve erken G 1’de eksprese edilen ve Sic 1 olarak adlandırılan inhibitörün bağlanması derhal engellenir. Çünkü Sic 1, özellikle B -tipi siklin-CDK komplekslerini baskılar ve S-faz inhibitörü olarak görev yapar
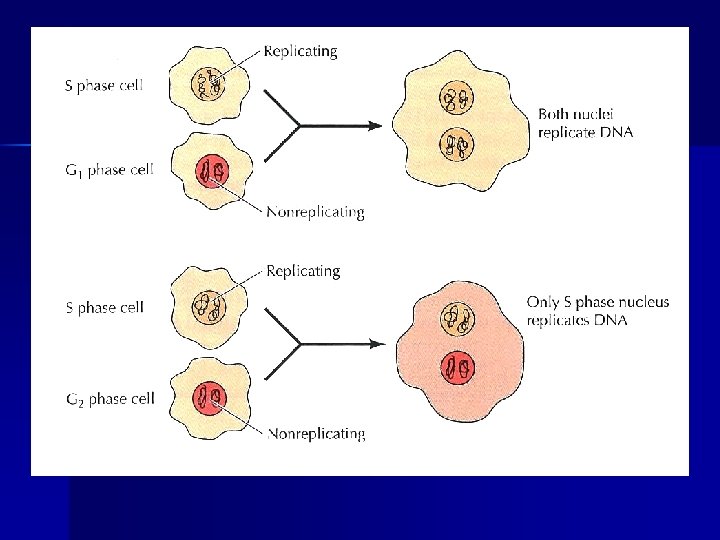
DNA replikasyonu erken G 1 boyunca orijinlerde toplanan replikasyon öncesi kompleksler tarafından başlatılır. n S-faz siklin-CDK kompleksleri replikasyonun eş zamanlı olarak replikasyon öncesi komplekslerden başlamasını tetikler ve yeni replikasyon öncesi komplekslerin oluşumu, replikasyon öncesi komplekslerin fosforlanmış olması nedeniyle yeniden bir araya gelmeleri engellenir n
TABLO 20 -2 Siklin-CDK Aktivitesinin Düzenleyicileri REGÜLATÖR TİPİ FONKSİYONU KİNAZLAR VE FOSFATAZLAR CAK kinaz Siklin-CDK’ların aktivasyonunu sağlar Wee kinaz Siklin-CDK’ları baskılar Cdc 25 fosfataz Siklin-CDK’ların aktivasyonunu sağlar Cdc 14 fosfataz Mitotik siklin-CDK’ları baskılamak için Cdh 1’i aktive eder Cdc 25 A fosfataz Omurgalı S-faz siklin-CDK aktivasyonunu sağlar Cdc 25 C fosfataz Omurgalı mitotik siklin-CDK aktivasyonunu sağlar ATM/ATR kinazlar Kontrol noktasını denetler, Chk 1/Chk 2 kinazları aktifleştirir Chk 1/Chk 2 kinazlar Kontrol noktasını denetler, hücre-döngüsü duraklamasına neden olan Cdc 25 C ve Cdc 25 A fosfatazların inaktivasyonunu sağlar BASKILAYICI PROTEİNLER Sic 1 S-faz siklin-CDK’lara bağlanıp baskılar CKI’lar p 27 KIP 1, p 57 KIP 2 ve p 21 CIP Siklin-CDK’lara bağlanıp baskılar INK 4 Orta-G 1 CDK’lara bağlanıp baskılar Mad 2 İğ-organizasyonu denetim noktasını kontrol eder, Cdc 20’ye bağlanarak anafazın başlamasını önler ve B-tipi siklin-CDK’ları inaktivasyonunu sağlar Rb E 2 F’lere bağlanır, birçok hücre döngü genlerinin transkripsiyonunu önler UBİKİTİN-PROTEİN LİGAZLAR SCF S-faz siklin-CDK’ları aktive etmek için fosforlanmış Sic 1 veya p 27 KIP 1’in yıkımını sağlar APC/C + Cdc 20 Sekürinin yıkımını uyarır, anafazı başlatır. B-tipi siklinlerin kısmi yıkımına neden olur APC/C + Cdh 1 Telofazı başlatmak için B-tipi siklinlerin tam olarak parçalanmasını uyarır ve metazoalardaki geminin, DNA replikasyon orijini üzerindeki replikasyon öncesi komplekslerin oluşmasını sağlar
TABLO 20 -3 Kontrol Nokta Proteinleri . KONTROL NOKTASI AMAÇ SENSÖR EYLEM S faz iç kontrol noktası M-faza girmeden önce tüm DNA replikasyonunun tamamlanmasını temin eder ATR replikasyon çatallarını tespit eder Mitotik siklin-CDK’ların aktivasyonunu önlemek için Cdc 25 C’nin baskılanması, erken mitotik olayların engellenmesi İğ-organizasyonu kontrol noktası Anafazdan önce tüm kromozom kinetokorlarının iğ mikrotübüllerine bağlandığından emin olur Mad 2 mikrotübüllere bağlanmamış kinetokorları tespit eder Separazın aktivasyonu için Cdc 20’nin baskılanması ve anafazın başlamasının önlenmesi İğ-pozisyonu kontrol noktası Telofaz ve sitokinezden önce tüm kromozomların yavru hücrelere eşit şekilde dağılmasını temin eder (S. cerevisiae) Tem-1 tomurcuktaki iğ kutup kitlesinin uygun pozisyonunu tespit eder Cdc 14 aktivasyonunun ve mitotik siklinlerin yıkımını ve geç mitotik olayların engellenmesi DNA-hasar kontrol noktası Hücre döngüsü boyunca DNA hasarını tespit eder ATM, ATR DNA hasarını tespit eder S faza girişi engellemek için Cdc 25 A’nın baskılanması, hücre döngüsü duraklamasını uyarmak için tüm siklin-CDK komplekslerinin p 21 CIP ile baskılanması
Hücre mitoza girebilmek için tüm genomunu kontrol eder, kromozom ve kromatid kırıkları gibi DNA hasarı varsa genomun gardiyanı olan p 53 sentezlenir ve hücre G 1 de bekletilir. Hasarın onarılması için süre verilir. Hasar onarılmazsa hücrede apoptosis denilen programlı hücre ölümü gerçekleştirilir.
Hücre döngüsünü baskılayan inhibitörler vardır. CİP/KİP(p 21, p 27, p 57) ailesi ve INK 4 ailesi(p 15, p 16, p 18, p 19)