Visprg mikrobioloija Mikroorganismu metabolisms Prof Uldis Kalnenieks Metabolisma

Vispārīgā mikrobioloģija: Mikroorganismu metabolisms Prof. Uldis Kalnenieks

Metabolisma iedalījums 1. Enerģijas avots Fototrofi Hemotrofi 2. Elektronu avots Litotrofi Organotrofi 3. Oglekļa avots Autotrofi Heterotrofi

Metabolisms: kopskats Izmantojamie substrāti un to katabolisma ceļi mikroorganismos ir ļoti dažādi; Būvbloku un biopolimēru biosintēze – samērā līdzīga
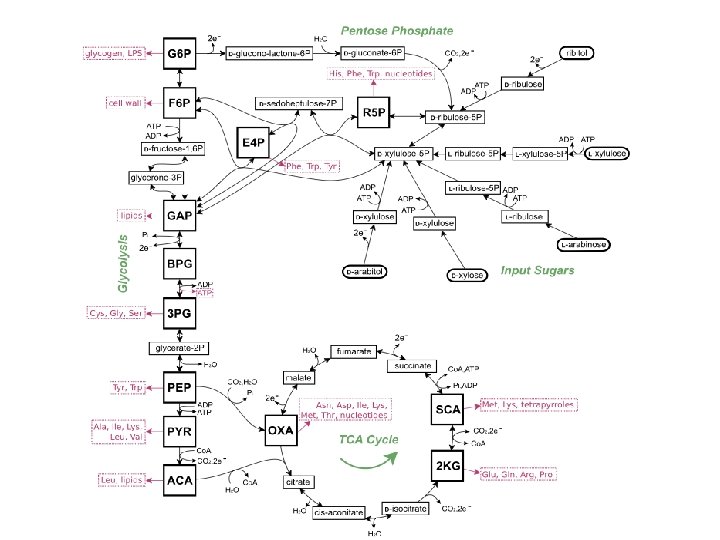

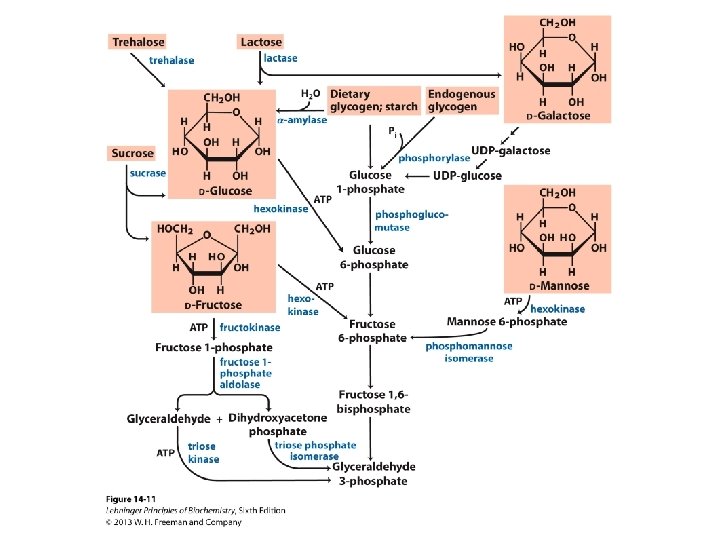
Mikroorganismu augšana uz ogļhidrātiem Polisaharīdi tiek degradēti ekstracellulāri un šūnā nonāk disaharīdi vai monosaharīdi Disaharīdi šūnā hidrolītiski vai fosforolītiski šķeļas par monosaharīdiem Monosaharīdi šūnā tiek fosforilēti, izomerizējas par glikozes vai fruktozes fosfātiem, kas tālāk nonāk centrālajos glikozes noārdīšanas ceļos

Pentožu cukuri pēc to transporta šūnā tiek fosforilēti, un pēc tam pentozofosfātu ceļa neoksidatīvā daļa kalpo pentožu cukuru pārveidošanai par fosforilētām heksozēm un/vai triozēm Pentožu efektīva izmantošana – mikroorganismu biotehnoloģijas problēma

Ogļhidrātu transportsistēmas: enerģētika un hierarhija PTS kontrolē pārējo transportsistēmu ekspresiju

Glukozo-6 -fosfāts Pentozofosfātu ceļš E. -M. -P. ceļš E. -D. ceļš

Entnera - Dudorova ceļa raksturīgās reakcijas Sastopams g. k. aerobajās Gramnegatīvajās baktērijās E. -D. ceļā veidojas tikai 1 ATP mols uz molu patērētās glikozes E. -D. ceļā tiek noārdīta arī glikonskābe (glukonāts) Tālāk sakrīt ar Embdena. Meijerhofa ceļa reakcijām pyruvate

plūsmu attiecību starp pentozofosfātu ceļu un glikolīzi ietekmē šūnas prasības pēc NADPH

Aerobs katabolisms ar pilnīgu ogļhidrātu oksidēšanos līdz CO 2 un H 2 O: piruvāts piruvātdehidrogenāzes komplekss Krebsa cikls elpošanas ķēde

Krebsa jeb trikarbonskābju jeb citronskābes cikls: - ir ogļhidrātu, aminoskābju un taukvielu oksidēšanās kopējais beigu posms; -tajā aktivētā acetilgrupa oksidējas līdz 2 CO 2 molekulām un veidojas 3 NAD(P)H, 1 FADH 2 un 1 ATP (vai GTP); NAD(P)H un FADH 2 oksidējas elpošanas ķēdē; - tā starpprodukti ir priekšteči aminoskābju u. c. biosintēzes reakcijām; - tā darbību inhibē, augstas ATP vai NAD(P)H koncentrācijas;

Daļai baktēriju Krebsa cikls ir nepilns, jeb ‘pakavveida’, un kalpo tikai biosintēzes vajadzībām
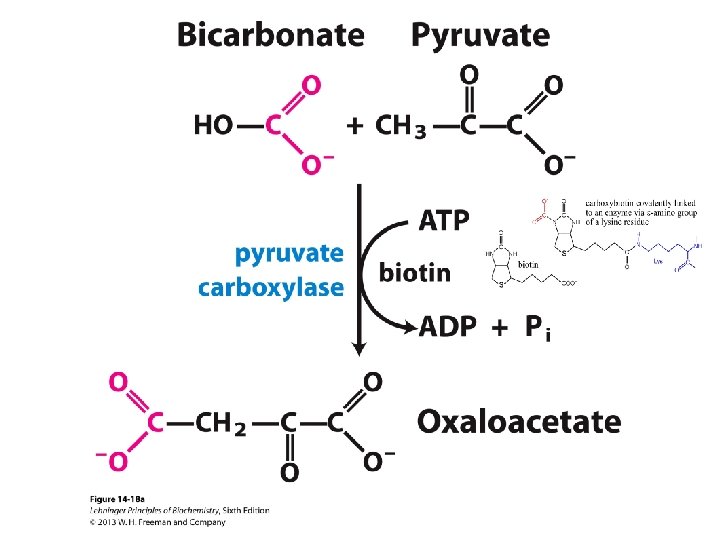
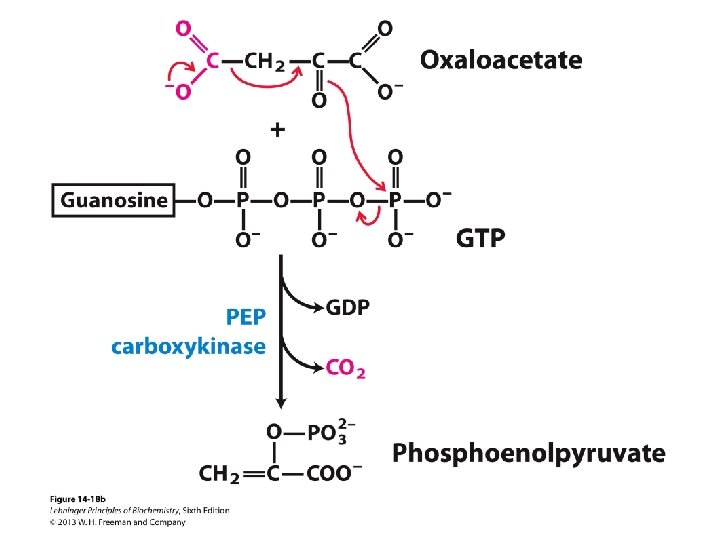
![Šūnai nepieciešamas īpašas anaplerotiskās jeb [centrālo metabolītu krājumus] papildinošās reakcijas, lai: - veidotu 3](http://slidetodoc.com/presentation_image_h/97ded9ef15f2f8904f80ec3e5cf31c85/image-15.jpg "Šūnai nepieciešamas īpašas anaplerotiskās jeb [centrālo metabolītu krājumus] papildinošās reakcijas, lai: - veidotu 3")
Šūnai nepieciešamas īpašas anaplerotiskās jeb [centrālo metabolītu krājumus] papildinošās reakcijas, lai: - veidotu 3 un vairāk C atomus saturošus centrālos metabolītus, ja augšana notiek uz substrāta, no kura veidojas acetil Co. A - papildinātu Krebsa cikla starpproduktu koncentrācijas, kuri tiek tērēti biosintēzēm

Vienu no 2 izocitrāta molekulām šķeļ izocitrātliāze 1 malāts papildus sintezējas no glioksilāta un acetil Co. A O=CH-COOH

acetil. Co. A: glioksilāta cikls + 5 pyr, lact: 3+1 vai 2+5 augot uz C 4 dikarbonskābēm cukuri: 1 vai 2
 J Bact 187, 1581 -1590 Ar zilu rāmīti atzīmēti centrālie")
Fuhrer et al. (2005) J Bact 187, 1581 -1590 Ar zilu rāmīti atzīmēti centrālie metabolīti – aminoskābju priekšteči Dažādu sugu baktērijām metaboliskie ceļi no centrālajiem metabolītiem līdz aminoskābēm ir vienādi, bet variē ceļš no glikozes līdz katram centrālajam metabolītam

Fluksomika – pēta plūsmu sadalījumu metaboliskajos ceļos dažādos kultūras augšanas apstākļos Eksperimentālajos pētījumos izmanto ar 13 C iezīmētus substrātus (piem. glikozi). Pastāv sakarība starp konkrētajiem metaboliskajiem ceļiem, pa kuriem veidojas metabolīts un iezīmētā oglekļa atomu sadalījumu tā molekulā; tas pēc tam viennozīmīgi atspoguļojas 13 C sadalījumā aminoskābēs: centrālo metabolītu biosintēzes ceļi “atstāj pēdas” aminoskābēs
 J Bact 187, 1581 -1590 Glikoze kā vienīgais oglekļa un")
Fuhrer et al. (2005) J Bact 187, 1581 -1590 Glikoze kā vienīgais oglekļa un enerģijas avots
 J Bact 187, 1581 -1590 Glikoze kā vienīgais oglekļa un")
Fuhrer et al. (2005) J Bact 187, 1581 -1590 Glikoze kā vienīgais oglekļa un enerģijas avots

Laboratorijas un industriālo modeļorganismu specifika: -EMP glikolīzes ceļš kā dominējošais; -pentozofosfātu ceļš kā viens no katabolisma ceļiem; -nepilnā oksidēšanās aerobajā katabolismā. Dabā plaši izplatīts ED ceļš!
- Slides: 24