LO SVILUPPO DEL FIORE FIORE complesso apparato di


")


 passa dalla")





 in Arabidopsis genera diversi organi in fasi diverse dello sviluppo")




")


")
 Identificazione di una rete")




 Scoperti in Drosophila, estremamente conservati negli Eucarioti In Drosophila determinano l’identità")
, AGAMOUS (arabidopsis), DEFICIENS (anthirrinum), SRF (uomo) In Arabidopsis oltre 100 geni")



, APETALA 2 (AP 2) Classe B:")
 specifica SEPALI ØA e B (AP 3, PI) specificano")












 induzione FPI ABCDE sviluppo FMI")

 AP 1 e CAL hanno sequenze strettamente correlate e un pattern di")






 (8 h luce;")












 CO promuove")
 m. RNA espresso nelle foglie m. RNA di FT e")


 + - evening genes luce +")
- Slides: 78
LO SVILUPPO DEL FIORE
ØFIORE: complesso apparato di strutture funzionalmente specializzate e radicalmente diverse dall’organismo vegetativo sia nella forma che nei tipi cellulari ØLa transizione verso la fioritura implica cambiamenti radicali nel destino delle cellule dei meristemi apicali ØAlterazione del destino delle cellule meristematiche: cambiamento di fase ØL’insieme degli eventi che portano l’apice del germoglio a produrre fiori si indica come INDUZIONE FIORALE
Struttura schematica di un fiore (ermafrodita)
Fiore staminifero Fiore pistillifero
INDUZIONE FIORALE Fattori endogeni: dimensioni, ritmi circadiani, ormoni Fattori ambientali: luce: fotoperiodo; temperatura: vernalizzazione Fotoperiodo: stimolo induttivo trasmissibile prodotto nelle foglie e recepito nel meristema (florigeno)
In Arabidopsis a seguito della INDUZIONE FIORALE Il meristema apicale vegetativo (MAV) passa dalla produzione ai suoi lati di foglie e primordi ascellari alla produzione di un numero indeterminato di PRIMORDI FIORALI Questo MERISTEMA APICALE si chiama MERISTEMA DELL’INFIORESCENZA Il MERISTEMA FIORALE si differenzia da un gruppo di cellule che si trovano ai lati del meristema dell’infiorescenza Il programma di sviluppo è diverso in quanto il meristema dell’infiorescenza è a crescita indeterminata mentre quello fiorale è a crescita DETERMINATA
ØMAV è convertito in meristema dell’infiorescenza in risposta ad uno stimolo fiorale ØIl meristema dell’infiorescenza è uno stato transitorio dalla crescita vegetiva a quella riproduttiva ØIl meristema dell’infiorescenza genera il MERISTEMA FIORALE
MERISTEMI FIORALI: distinguibili dai vegetativi perché di dimensioni maggiori Transizione allo stadio riproduttivo segnata da un aumento delle divisioni cellulari nella zona centrale del meristema apicale vegetativo AUMENTO DEL TASSO DI DIVISIONE DELLE CELLULE CENTRALI
Sezione longitudinale della regione apicale di un germoglio vegetativo e di un germoglio riproduttivo di Arabidopsis pf mg gf mg mf
Meristema vegetativo Meristema dell’infiorescenza
MERISTEMA DELL’ INFIORESCENZA E MERISTEMA FIORALE I primordi fiorali si originano nella stessa posizione nella quale si formano i primordi fogliari durante la crescita vegetativa; la fillotassi non è alterata
Il meristema apicale vegetativo(MAV) in Arabidopsis genera diversi organi in fasi diverse dello sviluppo All’inizio MAV forma una rosetta di foglie basali Nella fioritura MAV è trasformato in una infiorescenza primaria che produce un fusto allungato che porta le gemme fiorali
I primordi delle foglie formati prima della transizione alla fioritura diventano foglie caulinari, alla cui base si sviluppano infiorescenze secondarie le quali ripetono il programma di sviluppo dell’infiorescenza primaria
Infiorescenza indefinita Infiorescenza definita
Il meristema fiorale determina la formazione di quattro organizzati in cerchi concentrici verticilli La formazione degli organi più interni (carpelli) esaurisce le cellule meristematiche nella zona apicale e rimangono solo i primordi degli organi fiorali In Arabidopsis dall’esterno: ØQuattro sepali (verdi) ØQuattro petali (bianchi) ØSei stami (quattro più lunghi) ØUn pistillo (composto da due carpelli fusi a formare l’ovario; lo stilo e lo stigma)
Organi fiorali formati in sequenza dal meristema fiorale di Arabidopsis
Il fiore di arabidopsis (attinomorfo: simmetria raggiata)
Pistillo di Arabidopsis
Simmetria bilaterale Simmetria raggiata
Il fiore di Anthirrinum (zigomorfo: simmetria bilaterale)
Studi condotti principalmente su Arabidopsis e Anthirrinum (bocca di leone) Identificazione di una rete di geni che controlla la morfogenesi del fiore
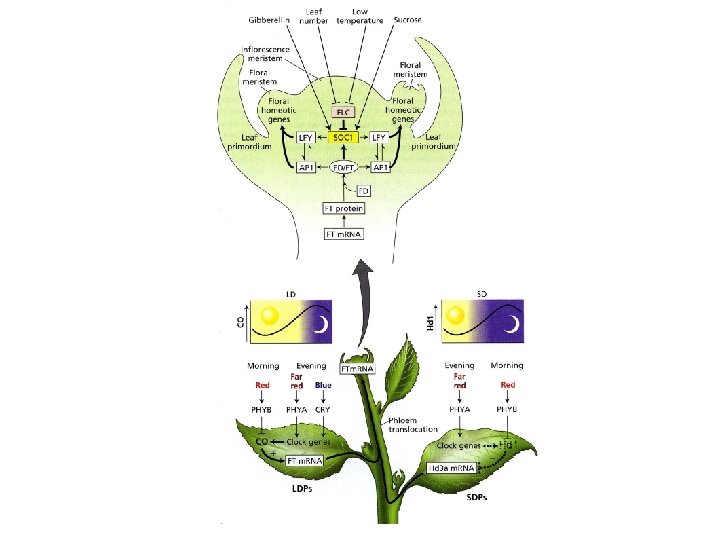
ØGeni che determinano l’identità degli organi fiorali (fattori di trascrizione che controllano l’espressione di geni di differenziamento degli organi del fiore; geni ABCDE ) ØGeni di identità meristematica (fattori di trascrizione necessari per la transizione da meristema vegetativo a fiorale e per la trascrizione dei geni di identità degli organi fiorali) In Arabidopsis SUPPRESSOR OF CONSTANS 1 (SOC 1); LEAFY (LFY); APETALA 1 (AP 1)
induzione FPI ABCDE sviluppo FMI
Lo studio di MUTANTI FIORALI OMEOTICI ha portato alla scoperta dei geni che controllano l’identità degli organi fiorali
I geni che determinano l’identità degli organi fiorali sono definiti OMEOTICI funzionalmente, in quanto mutazioni di questi geni comportano la formazione di un organo al posto di un altro (es: carpelli al posto di sepali). Tuttavia sono strutturalmente diversi dai geni omeotici animali in quanto non contengono il motivo HOMEOBOX ma appartengono alla classe di fattori MADS box è un motivo DNA binding conservato di 56 aa Inoltre è presente un motivo K box di interazione Proteina-proteina
GENI OMEOTICI (ANIMALI) Scoperti in Drosophila, estremamente conservati negli Eucarioti In Drosophila determinano l’identità anatomica dei segmenti Mutazione omeotica In Drosophila ANTENNAPEDIA Omeosi: trasformazione di una parte del corpo in un’altra
MADS: MCM 1(lievito), AGAMOUS (arabidopsis), DEFICIENS (anthirrinum), SRF (uomo) In Arabidopsis oltre 100 geni MADS BOX Animali o funghi ne possiedono un numero molto minore (Drosophila 2) Nelle piante molti coinvolti nello sviluppo del fiore APETALA 1, APETALA 3, PISTILLATA, AGAMOUS, SEPALLATA 1, SEPALLATA 2, SEPALLATA 3
MODELLO ABC Negli anni 90 E. Meyerowitz, E. Coen et al. proposero il modello ABC per l’identità degli organi fiorali ØBasato sullo studio dei mutanti fiorali omeotici di arabidopsis e anthirrinum Ømodello semplice e applicabile a molte angiosperme Øancora oggi valido con integrazioni
wt apetala 2 -2 pistillata 2 agamous 1
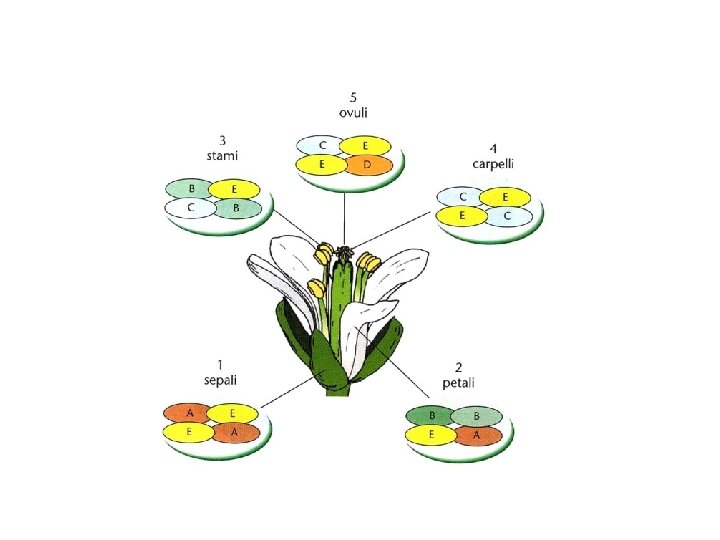
Il modello ABC postula che l’identità d’organo nel fiore è controllata dalla combinazione di tre funzioni geniche A B C nei verticilli fiorali A = SEPALI A+B = PETALI B+C = STAMI C = CARPELLI A e C si reprimono reciprocamente
In Arabidopsis Classe A: APETALA 1 (AP 1), APETALA 2 (AP 2) Classe B: APETALA 3 (AP 3), PISTILLATA (PI) Classe C: AGAMOUS (AG) In Anthirrinum Classe A: SQUAMOSA (SQUA) Classe B: DEFICIENS (DEF), GLOBOSA (GLO) Classe C: PLENA
ØA (AP 1, AP 2) specifica SEPALI ØA e B (AP 3, PI) specificano PETALI ØB e C (AG) specificano STAMI ØC (AG) specifica CARPELLI ØA e C si reprimono reciprocamente C (AGAMOUS) controlla anche lo stato di crescita determinata del meristema. L’assenza di C fa si che il meristema continui a crescere e nel quarto verticillo si formi un nuovo fiore
Fenotipi dei mutanti omeotici fiorali secondo il modello ABC wt agamous 1
apetala 2 pistillata 2
INTEGRAZIONE DEL MODELLO ABC Nel 1995 aggiunta la classe dei geni D Necessari a specificare l’identità dell’ovulo ABCD
INTEGRAZIONE DEL MODELLO ABCD Il mancato sviluppo di organi nelle posizioni corrette nei mutanti ABC dimostra che i geni ABC sono NECESSARI per specificare l’identità degli organi fiorali Sono anche sufficienti?
Nel 2000 scoperti i geni di classe E geni SEPALLATA MODELLO ABCDE geni SEP Øsono NECESSARI per specificare l’identità degli organi fiorali Øappartengono alla famiglia dei geni MADS
SEP 1, SEP 2, SEP 3, SEP 4. Sono simili nella sequenza e sono espressi in tutti i verticilli a partire dalla formazione del primordio fiorale
ØMutanti sep singoli o doppi non mostrano fenotipo ØTripli mutanti sep 1 sep 2 sep 3 esibiscono un fenotipo simile a doppi mutanti BC Come ad esempio pi ag oppure ap 3 ag (fiore formato da soli sepali) Nei tripli mutanti il fiore è formato interamente da sepali e i fiori sono indeterminati I geni SEP sono necessari per le funzioni dei geni di classe B e C
SEP 4 sep 1 sep 2 sep 3 : 1 verticillo sepali altri verticilli sepali omeotici (indeterminato) sep 1 sep 2 sep 3 sep 4: tutti i verticilli foglie omeotiche. Meristema fiorale indeterminato I geni sepallata sono necessari per la conversione delle foglie in organi fiorali
QUAL’E’ il meccanismo di interazione tra i geni ABC e i geni SEP ? Entrambi appartenenti alla classe MADS box e necessari per il corretto sviluppo degli organi fiorali Relazioni regolative a livello trascrizionale? (Ad esempio la trascrizione dei geni ABC può dipendere dai geni SEP o viceversa) NO SEP RNA si accumula in singoli mutanti ABC Espressione di ABC avviene nei tripli mutanti sep 1 sep 2 sep 3
Proteine MADS si legano in vitro al DNA come omo o etero dimeri INTERAZIONE DIRETTA A LIVELLO DELLE PROTEINE ABC /SEP ETERODIMERIZZAZIONE
MODELLO DEL QUARTETTO petali 2 eterodimeri che interagiscono tra loro
Il modello ABCDE e il modello del QUARTETTO
GENI DI IDENTITA’ MERISTEMATICA (FMI) induzione FPI ABCDE sviluppo FMI
GENI DI IDENTITA’ MERISTEMATICA fattori di trascrizione necessari per la transizione da meristema vegetativo a fiorale e per la trascrizione dei geni di identità degli organi fiorali In Arabidopsis: SUPPRESSOR OF CONSTANS 1 (SOC 1); LEAFY (LFY); APETALA 1 (AP 1) LEAFY è il principale interruttore tra i geni di identità meristematica: interrompe la crescita vegetativa e accelera la transizione alla fase riproduttiva
CAULIFLOWER (CAL) AP 1 e CAL hanno sequenze strettamente correlate e un pattern di espressione simile; all’inizio espressione in tutto il meristema, poi solo nel I e II verticillo del fiore Il gene CAULIFLOWER del cavolfiore coltivato ha un codone di stop che inattiva il prodotto genico Ciò previene la transizione dell’infiorescenza a meristema fiorale generando la ripetizione di meristemi dell’infiorescenza
L’ INDUZIONE DELLA FIORITURA
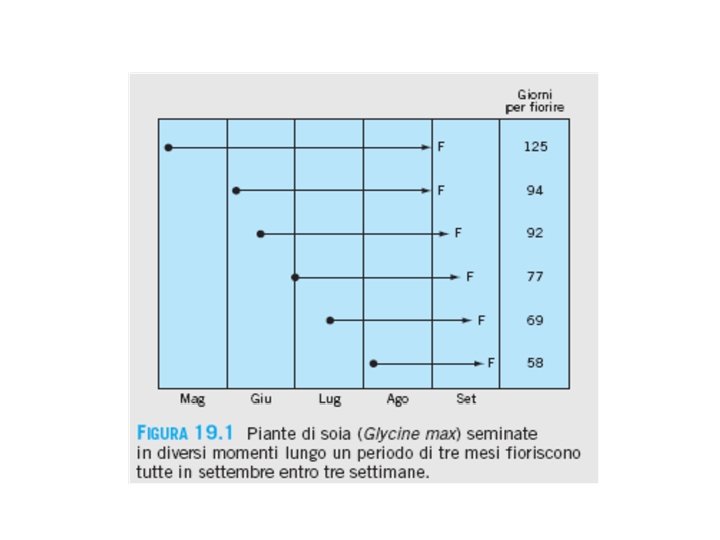
La fioritura e’ un fenomeno fotoperiodico il cambiamento della lunghezza del giorno può determinare l’inizio di un determinato processo fioritura
FOTOPERIODISMO: capacità di misurare la lunghezza del giorno per produrre una risposta fisiologica in un determinato momento dell’anno I ritmi circadiani e il fotoperiodismo hanno la proprietà comune di rispondere a cicli giorno/notte
Effetto della latitudine sulla durata del giorno La risposta fotoperiodica è influenzata dalla latitudine alla quale la pianta cresce
Le piante possono essere classificate secondo la loro risposta fotoperiodica rispetto alla fioritura
E’ CRITICA LA DURATA DEL PERIODO DI BUIO
Fioritura ritmica in risposta a interruzioni del buio nella SOIA (SDP) (8 h luce; 64 buio) Int 26 h= massima fioritura Int 40 h= fioritura nulla
DURANTE IL PERIODO DI BUIO SI ALTERNANO FASI DI SENSIBILITA’ E DI INSENSIBILITA’ ALLA LUCE • Nelle SDP (brevidiurne) la fioritura è indotta se l’interruzione del buio (alba) avviene dopo il completamento della fase sensibile alla luce • Nelle LDP (longidiurne) si ha fioritura se l’interruzione della notte coincide con la fase sensibile alla luce
La fioritura è un fenomeno fotoperiodico ed è fondamentale la misura del trascorrere del tempo durante il periodo di buio Ipotesi dell’ orologio (Bunning 1936) ØLa misura del tempo fotoperiodico si basa su di un ritmo circadiano in cui si alternano fasi di sensibilità e fasi di insensibilità alla luce ØLa capacità del segnale luminoso di indurre la fioritura dipende dalla fase del ritmo endogeno in cui il segnale viene recepito
Quando il segnale luminoso è percepito durante la fase sensibile del ritmo si può avere induzione della fioritura (LDP) inibizione della fioritura (SDP) SDP: fioritura se l’impulso è recepito DOPO il completamento della fase sensibile LDP: fioritura se l’impulso è recepito DURANTE la fase sensibile
MODELLO DELLA “COINCIDENZA ESTERNA” LPD: L’ESPOSIZIONE ALLA LUCE DEVE ESSERE COINCIDENTE CON LA FASE SENSIBILE DEL RITMO ENDOGENO
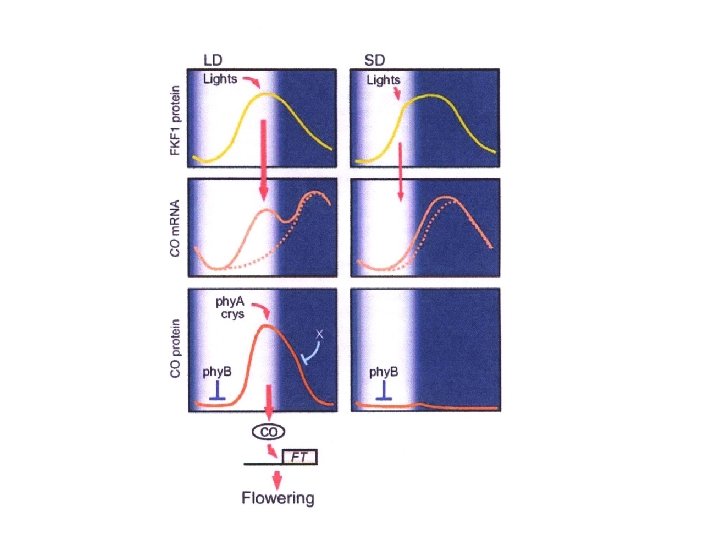
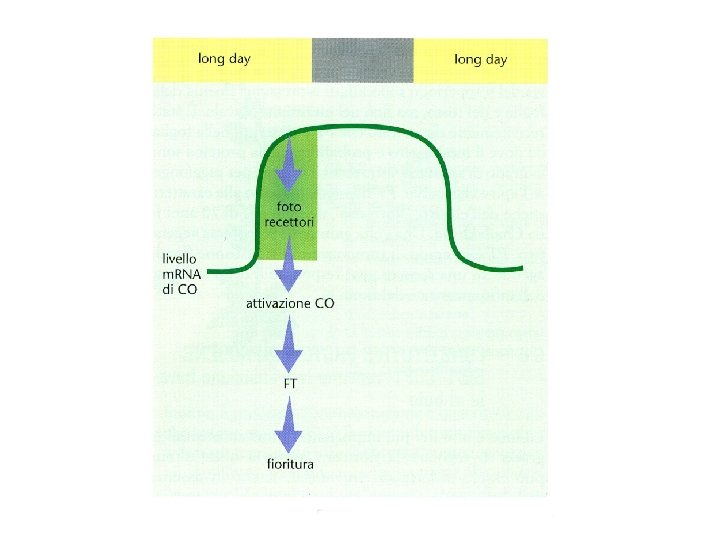
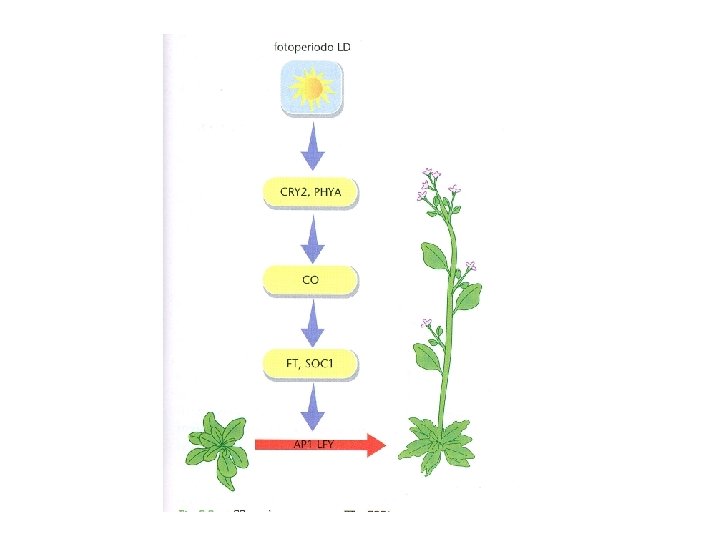
COINCIDENZA ESTERNA L’orologio circadiano genera una fase inducibile dalla luce durante il ciclo giorno/notte La fioritura si ha quando una particolare durata del giorno espone le piante alla luce durante la fase inducibile Individuati i componenti molecolari : L’interazione tra il ritmo circadiano e la luce avviene a livello della trascrizione del gene CONSTANS (CO) e della stabilità della proteina espressa CONSTANTS determina direttamente la trascrizione del gene FLOWERING LOCUS T (FT) che è un potente induttore della FIORITURA
Nel normale ciclo giorno/notte il m. RNA di CO mostra un ritmo giornaliero ØIn condizioni di GIORNO BREVE si hanno elevati livelli di CO m. RNA solo di notte ØIn condizioni di GIORNO LUNGOelevati livelli di CO m. RNa si raggiungono alla fine del periodo di luce
Deve esserci COINCIDENZA tra l’accumulo di m. RNA del gene CO e la luce del giorno In modo da consentire l’accumulo della proteina CO
Fioritura di Arabidopsis in condizioni di LD
Modello della COINCIDENZA ESTERNA La luce ha due funzioni: ØSincronizza l’orologio circadiano in modo da generare oscillazioni giornaliere di un regolatore chiave della fioritura con picco nel tardo pomeriggio Øregola l’attività di questo componente chiave
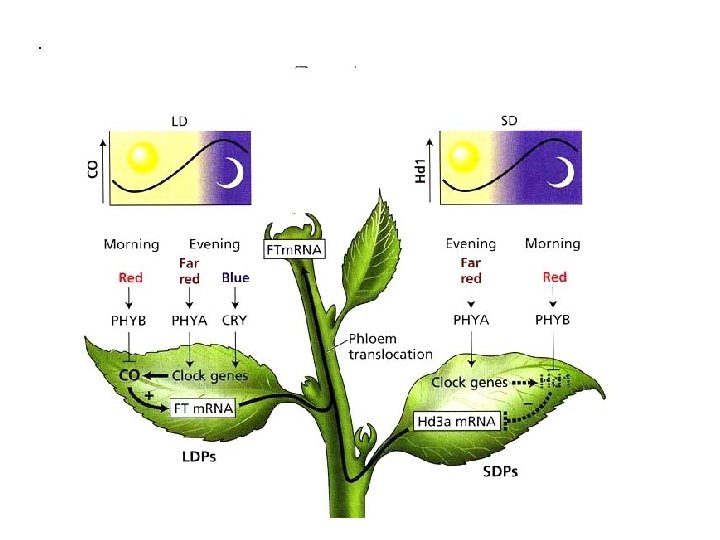
Come viene generato il ritmo giornaliero dei livelli della proteina CO? ØPhy. B promuove la riduzione dei livelli di CO al mattino ØPhy. A e criptocromi stabilizzano la proteina CO alla fine del giorno ØDurante la notte la proteina CO non stabilizzata è degradata
Induzione della fioriura in LPD e SPD Hd 1: ortologo di CO ma è un inibitore della fioritura Hd 3 a ortologo di FT
Il segnale fotoperiodico è percepito nelle foglie e trasmesso all’apice meristematico IPOTESI DEL “FLORIGENO”
CO è espresso nel sistema vascolare delle foglie (cellule compagne del floema) CO promuove fioritura se espresso nelle foglie ma non se espresso nel MAV
FT (FLOVERING LOCUS T) m. RNA espresso nelle foglie m. RNA di FT e la proteina possono migrare dalle foglie al MAV FT si lega al fattore di trascrizione FD espresso nel MAV Il complesso FT/FD attiva la trascrizione dei geni di identità meristematica (FMI) che determinano la transizione fiorale FT è il FLORIGENO
FT SOC 1 LEAFY FPI FMI LEAFY CAL AP 1 ABC
FINE
modello dell’oscillatore centrale in arabidopsis morning genes (LHCB) + - evening genes luce + TOC 1 + +