TRANSCRIPCIN EN TRIPANOSOMTIDOS 2007 Transcripcin en Tripanosomtidos 4

 •")
")
 y además, transcribe 2 grupos de")

 Nota: El ADN en el kinetoplasto está")
 • Es responsable de la transcripción de genes")








")



 • Glicoproteínas variables de superficie que constituyen una cubierta densa")



 VSG(interno) VSG(tel-act) VSG(copia-act) VSG(tel-inact) VSG(interno)")


- Slides: 29
TRANSCRIPCIÓN EN TRIPANOSOMÁTIDOS 2007
Transcripción en Tripanosomátidos • 4 ARN polimerasas (I, II y III; y mitocondrial) • Factor universal de transcripción relacionado con TBP (TRF 4: TBP related factor 4) • Organización general de genes: policistrónica (no relacionados funcionalmente) • “Capping” de ARNm (por trans-splicing) • La regulación post-transcripcional es la clave para los cambios en la expresión génica • Esta regulación se ejerce a través de elementos en las regiones intergénicas (rol de las regiones UTR)
ARN polimerasa III • Transcribe ARN pequeños (5 S, t. ARNs y sn. ARN) usando sn. ARN • Básicamente presenta los mismos promotores que eucariotas superiores: estar tanto en la región 3’ como 5’ Punto de inicio A Tipo 1 A Tipo 2 Tipo 3 ? ? C B ? ARNr 5 S ARNt U 1 -U 5 (diferencia con eucariotas sup)
ARN polimerasa I • Transcribe ARN ribosomal (nucleolo) y además, transcribe 2 grupos de genes que incluye Ags de superficie (VSG y PRO) • Los transcriptos ARNr no se le agrega cap: 20% 5’ tri-PO 4 y 80% 5’ OH • La estructura del promotor es semejante a los otros eucariotas: bipartita Punto de inicio – 170 – 150 – 130 – 110 Upstream control element – 60 – 40 – 20 – 10 +20 Core promoter I Pol
Promotores de ARN Pol I Similar a los promotores de ARNr euc. superiores Aparentemente están siempre activos (la regulación para las unidades de VSG y de PRO es en la elongación). Core Suficiente para transcribir VSG Se estimula la actividad por la presencia de elementos en el 5’ (UCE)
Mitocondrial transcripción: ARN polimerasa mitocondrial (mit. ARNpol) Nota: El ADN en el kinetoplasto está distribuído en: maxi- y mini- círculos 30 -50 kb 0. 8 -1. 6 kb 10 -30 copias 30000 -50000 copias genoma mitocondrial ARNguias No se han encontrado secuencias promotoras consenso mit. ARNpol es transcripta en el núcleo como una única subunidad Nota: los ARNt del kinetoplasto se transcriben en el núcleo y se deben importar
ARN polimerasa II (presenta dominio CTD) • Es responsable de la transcripción de genes estructurales (actina y tubulina), de los genes del miniexon (ARN-ME o SLRNA) y otros sn. ARN • Bajo nº de promotores y diferentes a otros eucariotas • No hay gran regulación en el inicio de la transcripción • Expresión constitutiva • Sin intrones (excep Poli(A) polim. de T. brucei, T. cruzi) • Transcripción policistrónica • la finalización de la transcripción al final del policistrón no está caracterizada aún • m. RNA maduro monocistrónico: capping, poli(A)
Promotores de ARN Pol II Como no existen evidencias de promotores clásicos de ARN pol II, se ha sugerido que inicia la transcripción al azar Se conoce: -Secuencias consenso en promotores para miniexon -Proteínas de unión a promotor del miniexon Los promotores se encuentran en el 5’ de cada copia (bipartita) -60 PBP 1 3 subunidades una TBP-like -30 PBP 2 I I l o P SL-RNA Evidencia que recluta la pol II T. . . T Finalización? (semejante a bacteria: rhoindependiente)
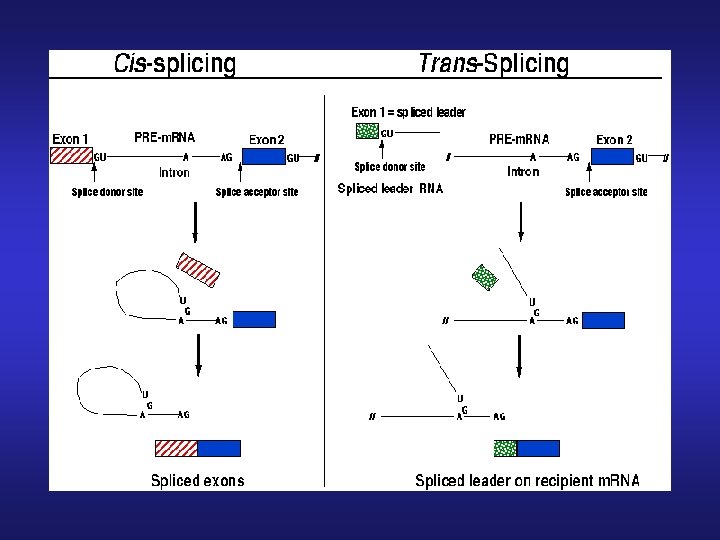
Procesamiento del pre-ARNm / Trans-splicing • Todos los ARNm estudiados comienzan con la misma secuencia de 30 -40 nt -> miniexon (ME - SL) • Esa secuencia no se encuentra en las vecindades del gen que está siendo analizado • Existen 100 -200 genes para ME por núcleo en tandem • c/u está presente en un repetido de aprox 1, 3 kb • Su transcripto presenta Cap 4 (m 7 G, metilaciones 2’OH ribosas 1 -4 y metilaciones en las bases 1 y 4) y es monocistrónico
El trans-splicing es un mecanismo utilizado para generar ARNm maduro para ser exportado del núcleo ADN
trans-splicing y poliadenilación están acoplados Trans-splicing alternativo: se describió para LYT una proteína de asociada con la virulencia. La diferncia son 28 aa en el N-t: La > es de membr (fase virulenta) la menor está en el kinetoplasto.
Transcripción de genes para proteínas es constitutiva No existen factores que regulen la iniciación de la transcripción Regulación se debe dar luego del inicio de la transcripción Elongación ME/poli(A) 3’UTR traduccional estabilidad de la proteína
Maquinaria de procesamiento de. ARNm transcripto policistrónico Podría ser más eficiente si se cambia la elección de un gen en un preexistente que en el ADN ya que se debe reclutar reguladores específicos (aunque esto implique una gran degradación del transcriptoma) ARN polimerasa encargada de la elongación Control sobre secuencias intergénicas
3’UTR de PRO: Estabilidad / Degradación Factores que regulan la producción de ARNm de PRO también regulan opuestamente el sitio activo de transcripción de la VSG ME 5’ UTR 27º citrato ME ME 3’ UTR AAAAA 37º no-citrato Transcripción basal AAAAA n n ME AAAAA ME n AAAA Se cree que estos factores externos actúan sobre elementos en el 3’UTR
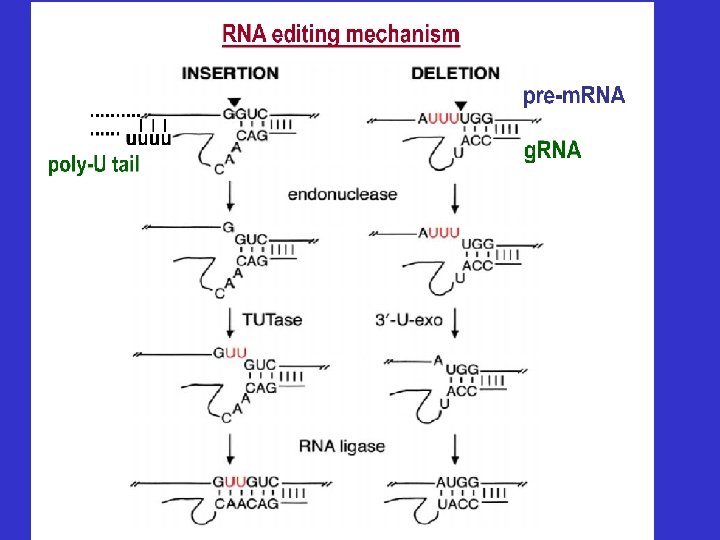
Editado • Modificación de la información génica codificada en el ADN. • Se adicionan o se remueven bases del ARNm. • Generalmente se introducen o se sacan “U” • Se corrige el marco de lectura • Ocurre en la mayor parte de los ARN mitocondrial. Simple: número pequeño de U en el extremo 5´ cox 2 en T. brucei Extensa: 60% del ARNm maduro no codificado por el gen > 50% de los genes de T. brucei mitocondriales, ej. Cox 3
Eucariotas inferiores: - En mitocondrias de Trypanosoma: la citocromo oxidasa sub. II (cox. II) que tiene insertadas 4 U que cambian aminoácidos y además cambia el marco de lectura. Como no existe otro gen, las U fueron agregadas luego de la transcripción - Mecanismo: se encontraron ARN-guías, que contiene una secuencia complementaria del transcripto Acción de la TUTasa usando pool de UTP de la célula Endorribonucleasa y ligasa (ATP)
CARACTERÍSTICAS DE LOS g. ARN • Todos los g. RNA tienen su propio gen en el minicírculos mitocondriales (salvo el 3’ de COXII) • Son pequeños (40 -70 nts) y no sufren edición. • El extremo 5’ de cada g. RNA es complementario a la secuencia cercana a la región editada hacia el 3’ del m. RNA. • El extremo 3’ de los g. RNAs tienen una cola de U (5 -15 nts), que es agregada por una terminal uridil transferasa (TUTasa). • La secuencia intermedia contiene una secuencia que es complementaria a la porción editada del m. RNA. (AU, GC y GU)
Algunas consideraciones del editado Armar marcos de lectura: - crear codón de inicio y de terminación, - establecer marcos de lecturas funcionales La edición es esencial para la diferenciación en T. brucei Las cantidades relativas de ARNm editado y sin editar varía en los diferentes estadios (mecanismo de regulación? )
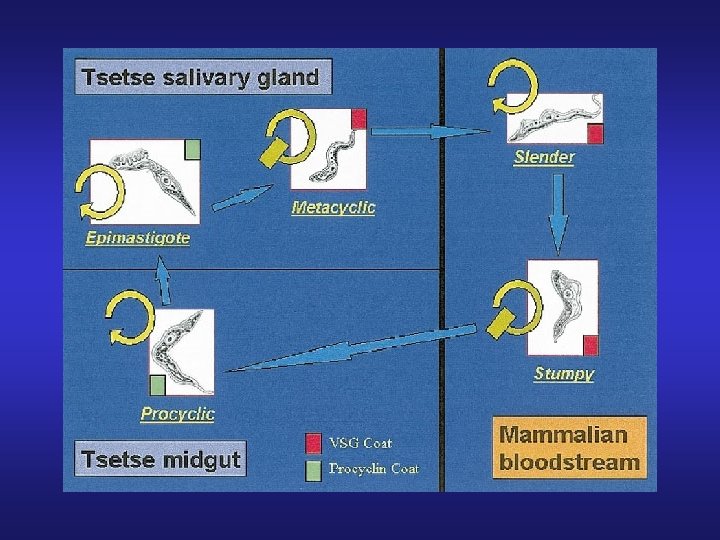
Variación antigénica T. brucei: Inmunofluorescencia de formas sanguíneas usando anticuerpos contra la VSG 221 (verde) o la VSG VO 2 (rojo).
Variant surface glycoprotein (VSG) • Glicoproteínas variables de superficie que constituyen una cubierta densa e inmunogénica de la membrana plasmática ( 107 por organismo) • Son grandes 450 -500 k. Da • Sólo una se expresa por vez • El cambio de expresión de VSG se realiza en forma espontánea (1 cada 100 por generación) • Existen aproximadamente 1000 potenciales variantes por organismo
• Los sitios de expresión están ligados a telómeros • Existen aproximadamente 20 sitios de expresión • El sitio activo de expresión aparentemente no se encuentra en un cuerpo nucleolar Sólo una se expresa, es decir que sólo se expresa la copia ELC que está localizada en el sitio de expresión que está bien cerca del telómero
• Cada cambio se acompaña de un pico de parasitemia Genes VSG: copia ligada a la expresión (ECL) 1. Southern blot: ADN de distintas poblaciones del parásito, Digestión con enzimas de restricción, Electroforesis en gel de agarosa, Transferencia a membrana de nylon, Hibridización. 2. Hirbridización con un ADNc de una población (pej. b) Copia Básica (BC) Copia ligada a la expresión (ECL) a b c d. . . etc Más del 50% de los genes VSGs siguen este patrón
Cada VSG es codificado por una copia básica que puede encontrarse a 5 -15 kb del telómero (teloméricos) o >50 kb del telómero (internos) VSG(tel-inact) VSG(interno) VSG(tel-act) Expresión algo programada (aparentemente involucra a la cantidad de repetidos de 70 pb)
La formación del ELC ocurre por conversión génica VSG(tel-inact) VSG(interno) VSG(tel-act) VSG(copia-act) VSG(tel-inact) VSG(interno) VSG(tel-act) VSG(interno) VSG(tel-inact)
Existen muchas copias del gen VSG en el genoma. La mayoría de los genes VSG son pseudogenes: recombinación puede generar nuevos genes VSG aumentando la variabilidad. Esta nueva información la guarda temporariamente cambiando el sitio de expresión Aparentemente el silenciamiento de los otros sitios de expresión y la recombinación son independientes
3000 pb VSG 1500 pb Región transpuesta 40 kb 5’ Repetidos cortos? 8 kb 3’ TELOMERO (CCCTAA)n Otro ejemplo es la bacteria Borrelia hermsii que causa la fiebre recurrente en el hombre. Esta bacteria sobrevive alterando una proteína de superficie (VMP), que están asociados con rearreglos en el genoma. El gen VMP activo está localizado cerca del telómero de un plásmido lineal.