CELL SINALLING TRANSDUCTION PATHWAYS TYPES OF SIGNALING MOLECULES


 direct")

 Extracellular molecules protein & peptides: Hormone, cytokine AA & its derivatives: Gly, Glu,")
 ENDOCRINE SIGNAL • • • Secreted signaling molecules or hormone by endocrine cells")
 Gaseous signal • • Simple structure, half life is short and active in")
 Intracellular molecules Ca 2+ ions • DG, ceramide IP 3 lipid derivatives carbohydrate")







 � Also known as seven-transmembrane domain receptors")






: The inactive form of PKA consist of two")









 protein a subunits, including;")





- Slides: 44
CELL SINALLING TRANSDUCTION PATHWAYS; TYPES OF SIGNALING MOLECULES AND RECEPTORS SOUMEN PATRA
1. INTRODUCTION � In human body, numerous processes are required for coordinating individual cells to support the body as a whole. � At the cellular level, Sensing of environments and cell communication for coordination relies on signal transduction; modeling signal transduction systems as self-organizing allows one to explain how equilibria are maintained. � Many disease processes, such as diabetes and heart disease arise from defects or dysregulations in these pathways, highlighting the importance of these processes in human biology and medicine
�Cell communication occurs through chemical signals and cellular receptors by either the 1) direct contact of molecules on two cells surfaces or the 2) release of a "chemical signal" recognized by another cell (near or far). Hormones are carried by the circulatory systems to many sites. Growth factors are released to act on nearby tissues. Ligands are signals that bind cell surface receptors (as observed with insulin (a ligand) and the insulin receptor) or that can pass into the cell and bind an internal receptor (such as the steroid hormones).
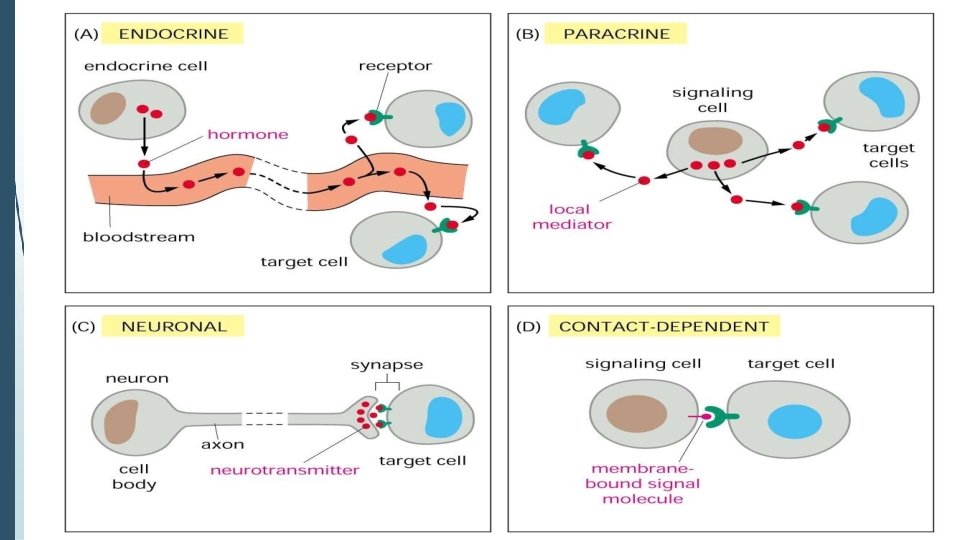
• Signaling molecules, which are released by signal-producing cells, reach and transfer biological signals to their target cells to initiate specific cellular responses.
1) Extracellular molecules protein & peptides: Hormone, cytokine AA & its derivatives: Gly, Glu, adrenaline, thyro xine Steroid: Sex Hormone, glucocorticosteroid Fatty acid derivatives: prostaglandin 1)Paracrine singnaling ( local chemical mediators) • Secreted signaling molecules act on nearby cells. • Reach neighboring target cells by passive diffusion. • Time of action is short. • Such as GF, PG
(2) ENDOCRINE SIGNAL • • • Secreted signaling molecules or hormone by endocrine cells and reach target cells by blood or lymph circulation to performed to their action. Time of action is long. Such as insulin, thyroxine, adrenalin 3) Synaptic signal (neurotransmitters) • • • Secreted signaling molecules (ions or neurotransmitters) by neuronal cells and reach another neuron by synaptic gap(synaptic cleft or gap junction) to generate action potential. Time of action is short. Such as Acetylcholine (Ach), noradrenaline, GABA etc.
4) Gaseous signal • • Simple structure, half life is short and active in chemistry. Such as NO, CO. 5) Autocrine signal • • Secreted signaling molecules act back to their own cells. Such as GF, cytokine, interferon, interleukin.
2) Intracellular molecules Ca 2+ ions • DG, ceramide IP 3 lipid derivatives carbohydrate derivatives • c. AMP nucleotides • Ras, JAK, Raf • c. GMP proteins
Small molecules synthesized in cells in response to an external signal are the second messengers, which are responsible for intracellular signal transduction. Such as Ca 2+, DG, Cer, IP 3, c. AMP, c. GMP Third messengers are the molecules which transmit message from outside to inside of nucleous or from inside to outside of nucleous, also called DNA binding protein.
� 2. SIGNAL TRANSDUCTION Any process occurring within cells that convert one kind of signal/stimulus into another type. It also known as cell signaling in which the transmission of molecular signals from a cell's exterior to its interior. Signals received by cells must be transmitted effectively into the cell to ensure an appropriate response. This step is initiated by cell-surface receptors which triggers a biochemical chain of events inside the cell, creating a response.
�It is also defined as the ability of a cell to change behavior in response to a receptor-ligand interaction. � The ligand is the primary messenger. � As the result of binding the receptor, other molecules or second messengers are produced within the target cell. � Second messengers relay the signal from one location to another (such as from plasma membrane to nucleus) leading to cascade of events/changes within a cell
SIGNAL TRANSDUCTION MECHANISMS 1/19/2016
�Messenger molecules may be amino acids, peptides, proteins, fatty acids, lipids, nucleosides or nucleotides. � Hydrophilic messengers bind to cell membrane receptors. � Hydrophobic messengers bind to intracellular receptors which regulate expression of specific genes. ´A ligand binds its receptor through a number of specific weak non-covalent bonds by fitting into a specific binding site or"pocket".
� 3. RECEPTORS � Receptors can be roughly divided into two major classes: intracellular receptors and extracellular receptors. � 4. EXTRACELLULAR RECEPTORS � Extracellular receptors are integral transmembrane proteins. � They span the plasma membrane of the cell, with one part of the receptor on the surface of the cell and the other on the cytoplasm. � Signal transduction occurs as a result of a ligand binding to the cell surface region of the receptor (Incase the ligand does not pass through the membrane).
� Various Extracellular Receptors � G protein-coupled receptors. � Receptors with Kinase activity. � Integrin receptors. � Toll gate receptors. � Ligand-gated ion channel receptors.
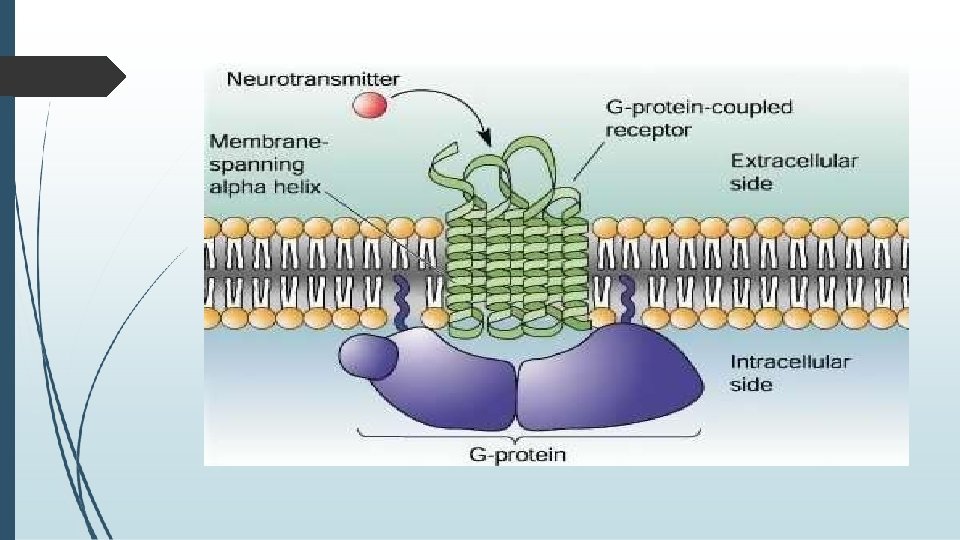
� 4 a. G PROTEIN–COUPLED RECEPTORS (GPCRs) � Also known as seven-transmembrane domain receptors (7 TM receptors) , heptahelical receptors, and G protein–linked receptors (GPLR). These constitute a large protein family of receptors that sense molecules outside the cell and activate several signal transduction pathways inside of a cell and ultimately create cellular responses. � The ligands that bind activate these receptors include � light-sensitive compounds � odors, pheromones, hormones, � and neurotransmitters, and vary in size from small molecules to peptides to large proteins.
�G proteins, also known as guanine nucleotide-binding proteins, are a family of proteins that act as molecular switches inside cells, and are involved in transmitting signals from a variety of stimuli outside a cell to its interior. � When they are bound to GTP, they are 'on', and, when they are bound to GDP, they are 'off'. � G proteins belong to the larger group of enzymes called GTPases
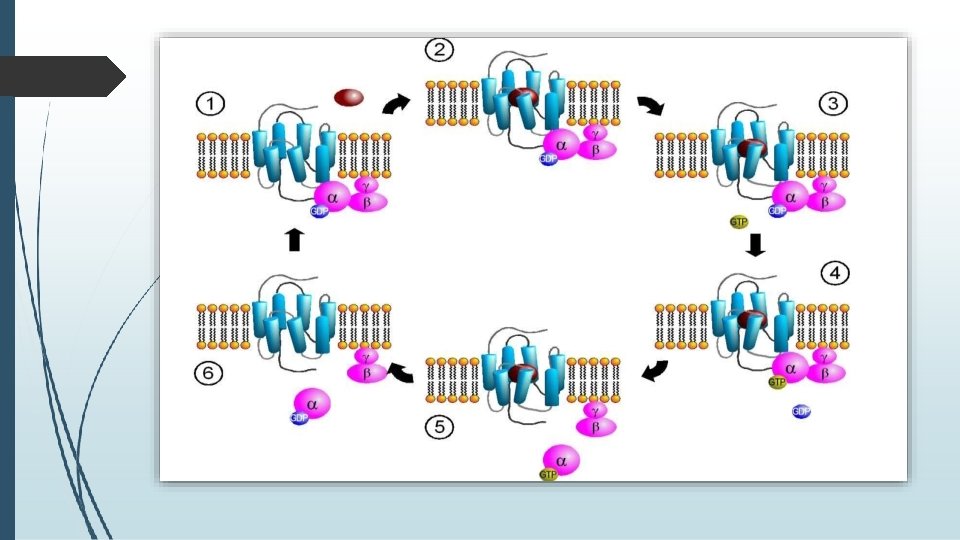
� There are two classes of G proteins. � The first function as monomeric small GTPases, � The second form and function as heterotrimeric G protein complexes. �Heterotrimeric class of complexes is made up of alpha (α), beta (β) and gamma (γ) subunits. The beta and gamma subunits can form a stable dimeric complex referred to as the beta-gamma complex while alpha subunit dissociates on activation.
� MECHANISM � It is known that in the inactive state, the GPCR is bound to a heterotrimeric G protein complex. � Binding of an agonist to the GPCR results in a conformation change in the receptor that is transmitted to the bound Gαsubunit of the heterotrimeric G protein. � The activated Gα subunit exchanges GTP in place of GDP which in turn triggers the dissociation of Gα subunit from the Gβγ dimer and from the receptor.
� The dissociated Gα and Gβγ subunits interact with other intracellular proteins to continue the signal transduction cascade. � While the freed GPCR is able to rebind to another heterotrimeric G protein to form a new complex that is ready to initiate another round of signal transduction.
�There are two principal signal transduction pathways involving the G protein–coupled receptors: � A. the c. AMP signal pathway and � B. the phosphatidylinositol signal pathway. � A. c. AMP-DEPENDENT PATHWAY, � It is also known as the adenylyl cyclase pathway. � In a c. AMP-dependent pathway, the activated Gs alpha subunit binds to and activates an enzyme called adenylyl cyclase, which, in turn, catalyzes the conversion of ATP into cyclic adenosine monophosphate (c. AMP).
�Increases in concentration of the second messenger c. AMP may lead to the activation of: �cyclic nucleotide-gated ion channels. � exchange proteins activated by c. AMP (EPAC) such as RAPGEF 3 � popeye domain containing proteins (Popdc) � and neurotransmitters, and vary in size from small molecules to peptides to large proteins.
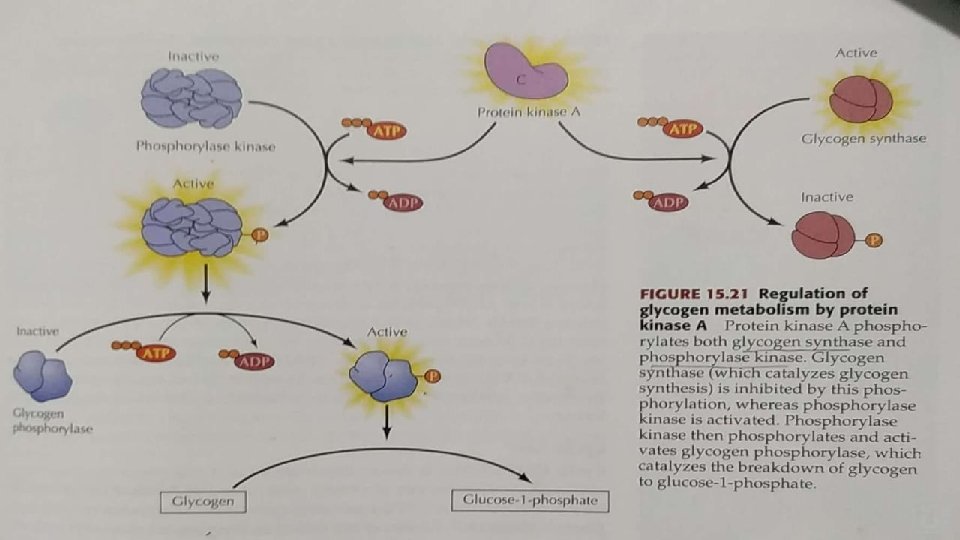
REGULATION OF PROTEIN KINASE A (PKA): The inactive form of PKA consist of two regulatory (R) and two catalytic (C) subunits. Binding of c. AMP to the R subunits induces a conformational change that leads to dissociation of the C subunits, which are then enzymatically active.
c. AMP-inducible gene expression: The free C subunit of PKA translocates to the nucleus and phosphorylates the transcription factor CREB(CRE-binding protein), leading to the recruitment of coactivators and expression of c. AMP-inducible genes.
� Molecules that activate c. AMP pathway include: � cholera toxin - increase c. AMP levels � caffeine and theophylline inhibit c. AMP phosphodiesterase, which degrades c. AMP - thus enabling higher levels of c. AMP than would otherwise be had. � pertussis toxin, which increase c. AMP levels by inhibiting Gi to its GDP (inactive) form. This leads to an increase in adenylyl cyclase activity, thereby increasing c. AMP levels, which can lead to an increase in insulin and therefore hypoglycemia
� DEACTIVATION � The Gs alpha subunit slowly catalyzes the hydrolysis of GTP to GDP, which in turn deactivates the Gs protein, shutting off the c. AMP pathway. � The pathway may also be deactivated downstream by directly inhibiting adenylyl cyclase or dephosphorylating the proteins phosphorylated by PKA. �Molecules that inhibit the c. AMP pathway include: � c. AMP phosphodiesterase dephosphorylates c. AMP into AMP, reducing the c. AMP levels � Gi protein, which is a G protein that inhibits adenylyl cyclase, reducing c. AMP levels.
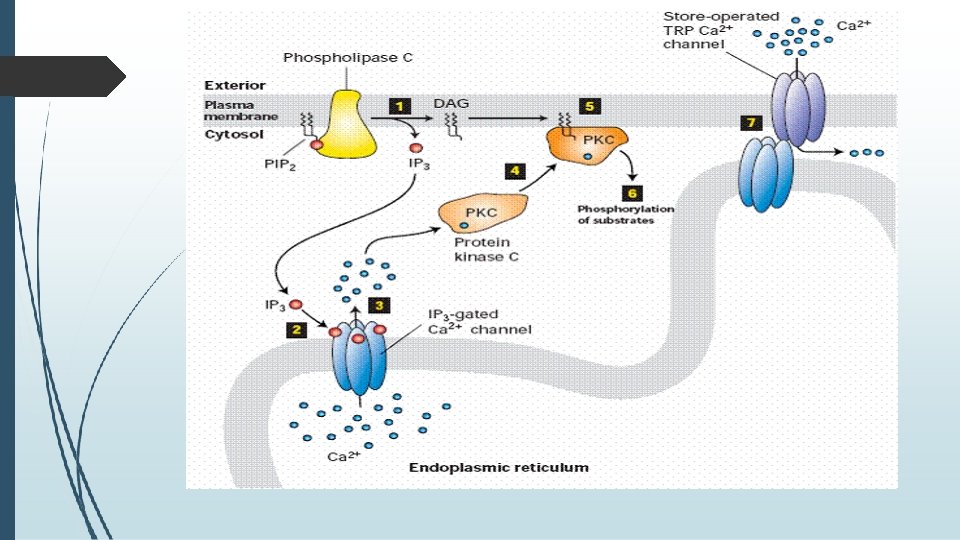
�B. PHOSPHATIDYLINOSITOL SIGNAL PATHWAY; �In the phosphatidylinositol signal pathway, the extracellular signal molecule binds with the G-protein receptor (Gq) on the cell surface and activates phospholipase C, which is located on the plasma membrane. phospholipase C
�IP 3 binds with the IP 3 receptor in the membrane of the smooth endoplasmic reticulum and mitochondria to open Ca 2+ channels. �DAG helps activate protein kinase C (PKC), which phosphorylates many other proteins, changing their catalytic activities, leading to cellular responses.
�The effects of Ca 2+ are also remarkable: � It cooperates with DAG in activating PKC and can activate the Ca. M kinase pathway, in which calcium-modulated protein calmodulin (Ca. M) binds Ca 2+, undergoes a change in conformation, and activates Ca. M kinase. � The kinase then phosphorylates target enzymes, regulating their activities. The two signal pathways are connected together by Ca 2+-Ca. M, which is also a regulatory subunit of adenylyl cyclase and phosphodiesterase in the c. AMP signal pathway.
� WHEN G PROTEIN SIGNALING IS DISRUPTED � G protein-related diseases are characterized by either deficient or excessive G protein signal transmission, which arises through abnormal signal initiation, defective termination, or reduced levels of G proteins � Deficient G protein signaling can arise through either reduced levels of G proteins, or through decrease signal initiation.
�Diseases involving a decrease in the production of G proteins include; � Night Blindness, where mutations in G(t) protein a subunits affect the response of rod cells to light. � Pseudo hypoparathyroidism, where the genetic loss of G(s) protein a subunits results in non-responsiveness to parathyroid hormone. � Other abnormalities involve decreased signal initiation through the inability of G proteins to switch to active states, e. g. the symptoms of Whooping Cough (Pertussis) like hypoglycemia
�Excessive G protein signaling can arise through either increased signal initiation, or defective signal termination. � Increased signal initiation occurs in Testotoxicosis, where a mutation in the receptor for luteinizing hormone can overstimulate G(s) proteins, resulting in the excessive production of testosterone. � Diseases arising from defective signal termination result from the persistent elevated activity of downstream effectors, such as in Cholera, the symptoms of which results from the action of a bacterial toxin that lead to stimulation of adenylyl cyclase and the subsequent secretion of salt and water leading to fatal diarrhea.
�Two other diseases involve defective termination through mutations in G(s) protein a subunits, including; � Adenomas, in which G proteins lose their ability to hydrolyze GTP through mutation, resulting in the excessive secretion of growth hormone and the increased proliferation of somatotrophs. � Mc. Cune-Albright Syndrome, where scattered regions of skin hyper-pigmentation arise from the hyper-functioning of one or more endocrine glands.
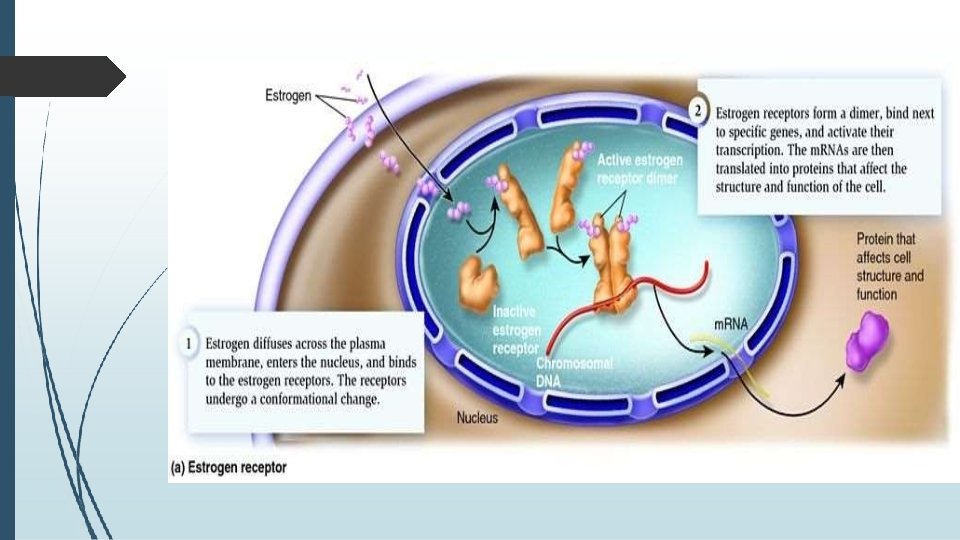
� 5. INTRACELLULAR RECEPTORS �Intracellular receptors are receptors located inside the cell rather than on its cell membrane. �Hormones that use intracellular receptors include thyroid and steroid hormones. �Examples are: Class of nuclear receptors located in the cell nucleus and cytoplasm and IP 3 receptor located on the endoplasmic reticulum.
� The ligands that bind to them are usually: � Intracellular second messengers like inositol trisphosphate (IP 3) � Extracellular lipophilic hormones like steroid hormones. � Activated nuclear receptors attach to the DNA at receptorspecific hormone-responsive element (HRE) sequences, located in the promoter region of the genes activated by the hormonereceptor complex. � Due to their enabling gene transcription, they are alternatively called inductors of gene expression.
�All hormones that act by regulation of gene expression have two consequences in their mechanism of action; � 1. their effects are produced after a characteristically long period of time � 2. their effects persist for another long period of time, even after their concentration has been reduced to zero, due to a relatively slow turnover of most enzymes and proteins that would either deactivate or terminate ligand binding onto the receptor. E. g. Thyroid Hormone
� CONCLUSION � So, the entire signal transduction system normally works astonishingly well, but sometimes serious problems can occur. � Mutations in growth factor receptors, G proteins, MAP kinases, and other molecules frequently contribute to cancer, and generally result in these molecules losing their normal switching function, staying in the activated form and therefore inappropriately stimulating these important enzyme cascades. � The complexity of the signaling system makes for challenging research, but once understood it holds the promise for better treatments for cancer and other diseases.