FOTOSINTEZA Sunevo zraenje svetlost obezbeuje toplotu energiju informacije

FOTOSINTEZA
 obezbeđuje: toplotu energiju informacije Fotobiologija")
Sunčevo zračenje (svetlost) obezbeđuje: toplotu energiju informacije Fotobiologija

Talasno-čestična priroda svetlosti Prostire se kao talas Oscilacije električnog i magnetnog polja Talasna dužina i frekvencija su obrnuto srazmerne Brzina prostiranja talasa Ponaša se kao čestica Foton - nosi kvant energije Energija kvanta zavisi od frekvencije Eλ h vac Monohromatska i polihromatska svetlost

Spektar elektromagnetnog zračenja Spektar Sunčevog zračenja Da bi došlo do hemijske promene svetlost mora biti apsorbovana

Kada dolazi do apsorpcije svetlosti Grotthuss-Draper-ov zakon -samo apsorbovana svetlost vodi hemijskoj promeni Stark-Einstein-ov zakon – 1 foton-1 molekul Apsorpcija fotona vodi prelazu elektrona u više energetsko stanje bitna je energija fotona a ne svetlosti

Atomska apsorpcija Molekulska apsorpcija U molekulu elektroni imaju veći “izbor” dostupnih energetskih nivoa jer su elektroni delokalizovani između dva jezgra A sistem konjugovanih dvogubih veza – delokalizacija π elektrona preko većeg broja jezgara – značajno povećava broj dostupnih energetskih stanja i smanjuje energetsku razliku između energetskih nivoa

Spektrofotometrija I T= Io A = - log T Io – intenzitet upadnog zraka I – intenzitet izlaznog zraka T – transparencija A – apsorbanca (ekstinkcija) Beer - Lambert – ov zakon : A=ε. c. l ε – molarni ekstinkcioni koeficijent c – koncentracija l – debljina stuba tečnosti

BOJA SUPSTANCE Apsorpcioni spektar

Apsorpcija u vidljivom delu spektra

Sudbina pobuđenog elektrona Viša pobuđena singletna stanja su veoma nestabilna Tripletna stanja su malo verovatna Prvi pobuđeni singlet ima četiri moguće sudbine: 1. 2. 3. 4. Termalna disipacija Fluorescencija Rezonantni transfer energije Fotohemija

ENERGETSKA STANJA HLOROFILA


Ključni eksperimenti u istraživanjima fotosinteze Engelmanov ogled Akcioni spektar fotosinteze Zelena rupa

Emerson i Arnold – koliko je molekula hlorofila potrebno za oslobađanje jednog molekula O 2 Kvantni prinos (0 -1) broj fotohemijskih produkata Ф= CO 2 + H 2 O broj apsorbovanih kvanata hν CH 2 O + O 2 467 k. J/mol Kvantna efikasnost (100 %) vs. energetska efikasnost (27 %)

Hillova reakcija Emersonov efekat pojačanja


1. Koncept fotosistema i prenosa energije 2. Razdvojenost faza fotosinteze
 i PS")
Na tilakoidnoj membrani se uočavaju globule supramolekulskih kompleksa PS II (P 680) i PS I (P 700) LHC I i II 20 -2000 molekula pigmenata CAB (Chl a/b vezujući prot. ) Cyt b 6/f 2 x. Cyt b, Cyt f, Fe-S centar ATP sintaza

Oksigena fotosinteza zahteva postojanje dva fotosistema Razdvojenost fotosistema implicira prisustvo mobilnih prenosioca elektrona i ukazuje da nije neophodna stehiometrija 1: 1 Mobilni prenosioci elektrona PQ, PC, Fd
 u proteinskim kompleksima – antena")
Rezonantni transfer energije Pigmenti organizovani u fotosisteme (karotenoidi, hlorofili) u proteinskim kompleksima – antena kompleks

Svi fotosistemi imaju zajedničko poreklo Antena kompleksi se veoma razlikuju

PSII reakcioni centar P 680 PSI reakcioni centar P 700
![Necikličan transport elektrona Redoks potencijal [V] Redoks potencijal Suština čuvanja energije u fotosintezi je](http://slidetodoc.com/presentation_image_h2/e5c75401ebbbe62a5862334f00547774/image-23.jpg "Necikličan transport elektrona Redoks potencijal [V] Redoks potencijal Suština čuvanja energije u fotosintezi je")
Necikličan transport elektrona Redoks potencijal [V] Redoks potencijal Suština čuvanja energije u fotosintezi je razdvajanje naelektrisanja u 1 ps (10 -12 s)
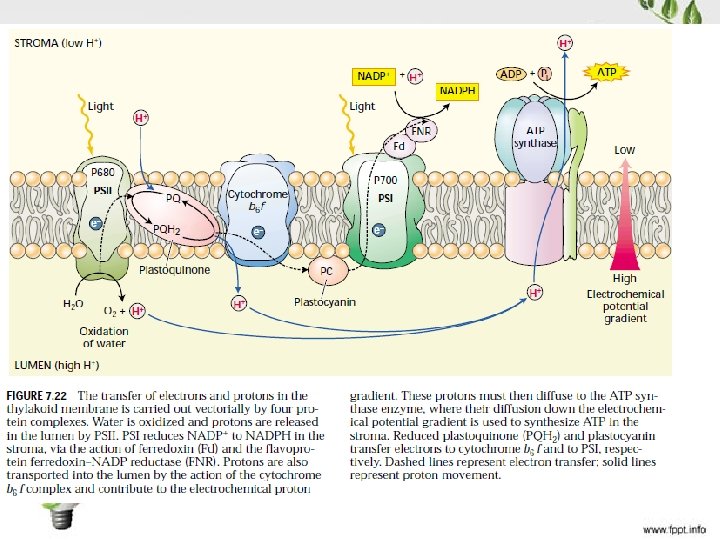

Transport elektrona oko PS II

Oslobađanje kiseonika

Transport elektrona na citohrom b 6/f kompleks Q-ciklus

Transport elektrona oko PS I

Inhibitori transporta elektrona DCMU Parakvat DBMIB Diuron, dibromotimokinon

Cikličan transport elektrona oko PS I Elektroni se sa Fd preko PQ na cyt b 6/f kompleks Nema oslobađanja O 2 niti redukcije NADP+, Jedini produkt je ATP

Fotofosforilacija Peter Mitchell 1960 -70, 1978 dobio Nobelovu nagradu Hemiosmotska hipoteza – kuplovanje sinteze ATP i transporta elektrona neophodnost istovremenog dešavanja in vivo Kuplujuća mesta i dekuplujući agensi (uncouplers) Elektrohemijski gradijent protona ∆ µH+= ∆p. H+ ∆ Ψm p. H gradijent između strome i lumena (Cl- i Mg++ kompenzuju električnu komponentu) Vektorski raspored redox komponenata (prenosioca elektrona) ATPaza u membrani, odnos H+/ATP
 Vektorski raspored redox komponenata (prenosioca elektrona) ATPaza")
Fotofosforilacija Kuplujuća mesta i dekuplujući agensi (uncouplers) Vektorski raspored redox komponenata (prenosioca elektrona) ATPaza u membrani, odnos H+: ATP 4: 1

Mehanizam sinteze ATPa Prolazak protona Rotacija γ subjedinice Promena konformacije vezujućeg mesta na αβ dimerima O- open (otvoreno) L - loose (labavo) T - tight (tesno)

Jedan od najubedljivijih dokaza za hemiosmotsku hipotezu bila je tzv kiselo-bazna fosforilacija In vivo je neophodno funkcionisanje oba procesa

Usklađivanje sinteze ATP i NADPH Kada je pul adenilata pretežno redukovan, sistem je zasićen elektronima, što zaustavlja i sintezu ATPa. Zato postoji ciklični transport elektrona oko PSI U regulaciji učestvuje i LHCII, koji zavisno od potrebe može biti vezan za PS II ili PS I

Pseudocikličan transport elektrona • U odsustvu akceptora, kada je pul feredoksina jako redukovan, dolazi do Mehler-ove reakcije, transfera elektrona na molekulski kiseonik pri čemu se generiše superoksid anjon radikal.

Fotoinhibicija fotosinteze – generisanje ROS Fe 3+ + • O 2− → Fe 2+ + O 2 Fe 2+ + H 2 O 2 → Fe 3+ + OH− + • OH Fenton • O 2 - + H 2 O 2 → • OH + OH- + O 2 Haber-Weiss
 Askorbat-glutationski ciklus (Halliwel-Asada ciklus) Regeneracija Asc i")
Ksantofilni ciklus “gašenje” pobuđenog stanja hlorofila (quenching) Askorbat-glutationski ciklus (Halliwel-Asada ciklus) Regeneracija Asc i potrošnja NADPH Ciklus voda-voda Uklanjanje ROS

Fotosinteza je ograničena koncentracijom CO 2 i intenzitetom svetlosti. Temperatura utiče samo na deo fotosinteze koji zavisi od CO 2 dok nema uticaj na deo koji zavisi od svetlosti

faze fotositeze svetla i tamna fotohemijska i biohemijska • Fotohemijska faza podrazumeva • Apsorpciju svetlosti i oksidaciju hlorofila u RC • Redukciju H 2 O i oslobađanje O 2 • Kretanje elektrona kroz redoks lanac • Redukciju NADP+ • Sintezu ATPa

Biohemijska faza fotosinteze Koriste se ATP i NADPH sintetisani tokom fotohemijske faze
 Prvi produkt nije saharoza")
Berkley, Amerika 40 -ih i uvođenje radioaktivnih izotopa (14 C) Prvi produkt nije saharoza Prvi stabilan intermedijer je 3 -fosfoglicerat Brzo se javlja još veliki broj jedinjenja
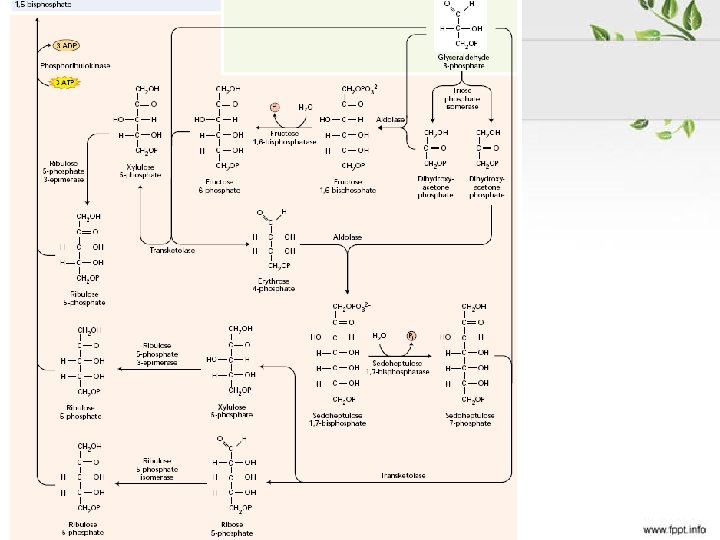
 Karboksilacija – vezivanje CO 2 u organsku materiju, za ribulozo-1,")
Kalvinov ciklus (reduktivni PPC) Karboksilacija – vezivanje CO 2 u organsku materiju, za ribulozo-1, 5 -bisfosfat Redukcija – od oksidovanog (C+4) do redukovanog (C+1) u GAP Regeneracija akceptora – Ru-1, 5 -b. P

Rubis. CO • Jedini enzim koji omogućuje fiksaciju atmosferskog CO 2 za izgradnju biomase • Katalizuje i oksigenaciju • Do 40% solubilnih proteina lista • Najzastupljeniji protein na Zemlji • Afinitet prema CO 2 je dovoljno visok da enzim radi i pri niskim koncentracijama CO 2

• Fosforilacija fosfoglicerata • Redukcija bisfosfoglicerata • NADPH GA-3 -P dehidrogenaza • Enzim sličan citosolnom koji učestvuje u glikolizi

Da bi tekao ciklus neophodna je regeneracija akceptora – Ru. BP Samo 1/6 triozo fosfata se koristi za sintezu skroba i saharoze Brzina ciklusa zavisi od koncentracije intermedijera (lag faza pri uključivanju osvetljenja)
- Slides: 47