Antigeni Lantigene una macromolecola capace di reagire con

 Flagelli (proteine) Genoma")








 comporta il")
 di classe Il linfocita B maturo in seguito al contatto con l’antigene")
- Slides: 13
Antigeni L'antigene è una macromolecola capace di reagire con i prodotti del sistema immunitario. Viene definita antigene ogni qualsivoglia sostanza non appartenente a un dato organismo (non-self). Gli antigeni sono solitamente di natura proteica, ma possono essere di qualsiasi natura chimica, come polisaccaridi, o acidi nucleici. Nella definizione di antigene bisogna distinguere due caratteristiche fondamentali: – l'antigenicità (specificità antigenica) rappresenta la capacità dell'antigene di combinarsi specificamente con anticorpi e recettori per l'antigene; – l'immunogenicità o potere "immunogeno" indica la capacità dell'antigene di indurre una risposta immunitaria. • La piccola parte di antigene che lega l'anticorpo specifico è detta epitopo (o determinante antigenico). La singola molecola di antigene può contenere diversi epitopi riconosciuti da anticorpi differenti
Localizzazione dei principali antigeni microbici Antigeni batterici Membrana cellulare (proteine, glicoproteine) Flagelli (proteine) Genoma (DNA) Citoplasma (proteine, polisaccaridi, RNA) Pili e fimbrie (proteine) Capsula (polisaccaridi) Parete (acidi teicoici) Membrana esterna (gram-) (polisaccaride) Altri antigeni batterici: porine (proteine); esotossine, etc Antigeni virali: (1) glicoproteine del pericapside; (2) proteine del tegumento; (3) proteine del capside; (3) proteine precoci e tardive; (4) m. RNA; (5) RNA e DNA genomici.
Rapporto fra antigenicità, immunogenicità e costituzione chimica L’immunogenicità di una molecola dipende da diverse caratteristiche, come: (1) la natura chimica; (2) la dimensione; (3) l’estraneità all’organismo (i nostri antigeni sono definiti self, quelli estranei non self); (4) la via di entrata dell’antigene, ad esempio via orale, via inalatoria, via cutanea, via parenterale (intramuscolare, endovenosa). Gli apteni sono molecole piccole che non sono dotate di immunogenicità, diventano immunogene quando sono legate a una molecola proteica, chiamata carrier.
Antigeni proteici o peptidici Epitopi sequenziali Determinanti conformazionali Determinanti sequenziali epitopi o determinanti sequenziali, caratterizzati da una specifica sequenza lineare aminoacidica (ad esempio Cys-Gly. Ser-His-Leu-Val-Glu-Ala-Leu-Tyr) epitopi o determinanti conformazionali, riconosciuti dal sistema immunitario come complessi tridimensionali. Gli epitopi conformazionali possono essere costituiti da elementi anche molto distanti tra loro in termini di sequenza primaria (lineare), ma estremamente vicini a livello della struttura terziaria e quaternaria (tridimensionale) a causa del ripiegamento che caratterizza molte macromolecole biologiche.
Molecole in grado di legare l’antigene MHC-I lega peptidi di 8 -11 aa; MHC II lega peptidi di 30 o più aa Trasduzione del segnale BCR: Recettore delle cellule B formato da immunoglobuline di membrana: Ig. M e Ig. D nei linfociti B naive, che non hanno ancora incontrato l’antigene e da due molecole di trasduzione del segnale, Igα e Igβ (CD 79). TCR: Recettore delle cellule T riconosce l’epitopo antigenico associato all’MHC ed è associato alle molecole CD 3 e zeta per la trasduzione del segnale TCR BCR
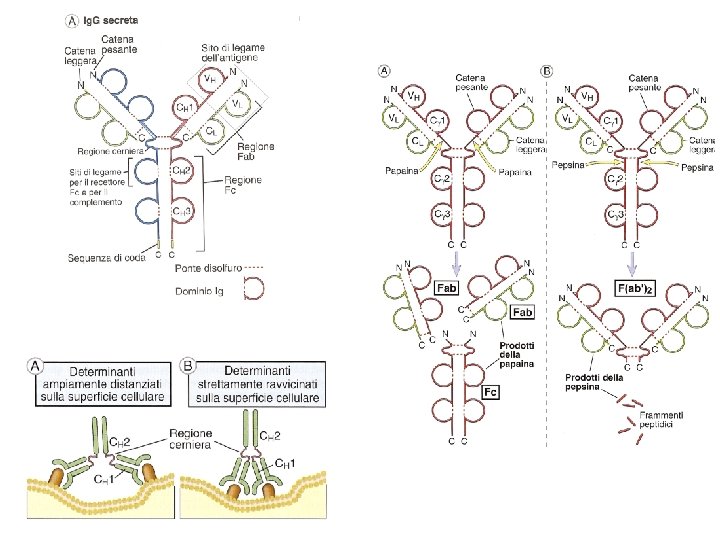
Struttura e caratteristiche di una immunogloblina CDR 1 CDR 2 CDR 3 Regioni ipervariabili= CDR 1 -3 (complementary determining regions)
Classi anticorpali
Ig. A secretorie
Origine della diversità recettoriale Riarrangiamento dei geni del recettore per l’antigene nei linfociti B e T • I geni che codificano per i recettori per l’antigene sono composti da differenti segmenti genici che codificano per la regione variabile (V), per la diversità (D) e/o per la ricongiunzione ( joining-J). • Nei linfociti B, che maturano nel midollo osseo, i geni che codificano per le catene leggere ( e ) si trovano rispettivamente sui cromosomi 2 e 22, mentre il gene che codifica per la catena pesante si trova sul cromosoma 14 • Nei linfociti T, che maturano nel timo, i geni per le catene e si trovano sul cromosoma 7, mentre i geni per le catene e si trovano sul cromosoma 14.
Stadi della maturazione dei linfociti B • I linfociti B e T originano da un precursore comune di derivazione midollare dove, in seguito, la maturazione dei B avviene nel midollo osseo mentre i T migrano al timo dove completano la maturazione • I progenitori delle cellule B passano attraverso diversi stadi di differenziazione dando origine alle cellule B immature che esprimono molecole Ig. M sulla membrana. • I linfociti B immaturi lasciano il midollo per continuare la loro maturazione nella milza, dove le cellule B mature follicolari esprimono sulla membrana Ig. M e Ig. D • I cloni linfocitari maturano e acquisiscono la loro specificità, in assenza dell’antigene, negli organi linfoidi primari (midollo osseo per i linfociti B e timo per i linfociti T) e nel passaggio alla milza. I cloni di linfociti maturi specifici per i diversi determinantigenici entrano nei tessuti linfoidi secondari dove vengono attivati dall’incontro con l’antigene e si differenziano in (1) cellule effettrici (es. plasmacellule che secernono anticorpi) e (2) linfociti della memoria.
Origine della diversità anticorpale La maturazione dei linfociti B (in assenza dell’antigene) comporta il riarrangiamento somatico dei diversi segmenti genici, V, J per la parte variabile delle catene leggere K e e V, D, J per la parte variabile delle catene pesanti. Nell’uomo esistono circa 35 geni V per la catena leggera e 100 segmenti genici V per la catena pesante. Origine delle differenze anticorpali: 1. Segmenti genici multipli per le regioni variabili VDJ 2. Unione casuale dei segmenti VJ sulle catene leggere o VDJ sulle catene pesanti 3. Associazione di una qualsiasi catena leggera con una qualsiasi catena pesante. Questo meccanismo origina 6, 4 x 106 diversità anticorpali Altri 3 meccanismi portano le differenze a più di 109 diversità. 4. Diversità al punto di unione VJ o VDJ 5. Addizione di nucleotidi arbitrari a livello della giuntura VDJ 6. Ipermutazione somatica dei geni riarrangiati
Commutazione (cambiamento) di classe Il linfocita B maturo in seguito al contatto con l’antigene prolifera e si differenzia in plasmacellula e linfociti B della memoria. La differenziazione in plasmacellula che secerne specifiche classi anticorpali dipende quasi sempre (ad eccezione delle Ig. M) da un meccanismo chiamato “commutazione di classe”, che è antigene timo-dipendente e pertanto necessita dell’aiuto di un linfocita TH. La classe dell’anticorpo dipende dal tipo (1) di antigene, (2) di TH e (3) di citochina. Il cambiamento di classe da Ig. M a Ig. G o Ig. E o Ig. A viene indotto nei linfociti B maturi da citochine prodotte dai linfociti T helper. Ciascun segmento genico è preceduto da una sequenza di DNA detta sequenza switch (S). Ad esempio, il segnale di specifiche citochine induce la ricombinazione della sequenza S che precede la sequenza μ con la sequenza S che precede le sequenze ; creando un’ansa che verrà rimossa. Il processamento dell’RNA genera il messaggero definitivo per la catena delle immuoglobuline. Lo scambio di classe può avvenire attraverso due meccanismi: (A) la delezione di alcuni segmenti genici C (es. , , ) e la ricombinazione del complesso VDJ con il gene C successivo (es. e vedi figura). (B) la processazione (splicing) alternativa di un trascritto primario (es. ) Iα Cα