Uren pohlav a pohlavn chromozomy sexualita z genetickho

")

 chromozom s 1, 2, 3. .")

 Degeneraci nepárového pohlavního chromozomu doprovází inaktivace pohlavních chromozomů v meiotickém dělení")


 Homologie s chromozomy ZW")
 Homologie s chromozomy ZW")



 nejsou inaktivovány. inaktivace je řízena z tzv. inaktivačního")

 z neoplozených vajíček")


 fungují odlišně")
- Slides: 20
Určení pohlaví a pohlavní chromozomy sexualita - z genetického hlediska střídání n a 2 n v životním cyklu evolučně původní je asexualita, původním typem sexuality je hermafroditismus význam sexuality: rozrůznění genotypů Nejen hermafroditi, ale i gonochoristé potenciálně bisexuální. U hermafroditů rozhodujií o realizaci maskulinizačních nebo feminizačních faktorů vlivy vnějšího či vnitřního prostředí. Proterandie, proterogynie. Dědičně fixována doba, kdy jsou organismy k těmto vlivům citlivé (embryonální vývoj - Sagitta, larvální vývoj - Bonelia, adulti - mnohoštětinatci). Jedná se o tzv. epigamní (metagamní) určení pohlaví. Extrémní případ epigamie: určení pohlaví vnějšími vlivy v průběhu oogeneze (tzv. progamní určení pohlaví). simultánní hermafroditi: poloha buněk v těle (nezmar) sukcedánní hermafroditi: vývojové stáří, velikost jedince atp. : plž Crepidula (řetězce jedinců: spodní ♀, nad nimi intesexy, nahoře ♂) fenotypový gonochorismus: hermafroditismus již rudimentární, přesto vnější vlivy stále určují pohlaví: Bonelia, mořské želvy U typických gonochoristů určení pohlaví pri oplození vajíčka (syngamně), relativně nezávisle na prostředí.
Mechanismus: segregace a kombinace maskulinizačních a feminizačních faktorů nesených pohlavními chromozomy (= gonozomy, heterochromozomy) (sex chromosomes) Objev: * Henking 1891, Mc. Clung 1902, Wilson 1905 U typických gonochoristů poměr pohlaví 1: 1. . . stejně jako při křížení Aa x aa Tzn. jedno pohlaví analogické heterozygotovi (heterogametické), druhé recesivnímu homozygotovi (homogametické). Význam poměru 1: 1: při křížení: poskytuje maximální počet možných kombinací (tedy max. genotypovou variabilitu), zároveň se snižuje možnost příbuzenského křížení Nepárový chromozom heterogametického pohlaví - alozom Dva typy heterogametie: samčí - typ Drosophila podtypy: Lygaeus (XY/XX), Protenor (X 0/XX) samičí - typ Abraxas (méně častý), ZW/ZZ Výskyt jednotlivých typů heterogametie: Drosophila - vrtejši, hlístice, většina skupin hmyzu a pavoukovců, ostnokožci, některé ryby, většina bezocasých obojživelníků, želvy, savci rostliny: mechorosty, Rumex, Humulus, Cannabis, Papaya, Mercurialis, silenkovité Abraxas - motolice čel. Schistosomatidae, řada korýšů, motýli + chrostíci, některé ryby, ocasatí obojživelníci, řada plazů (zejm. šupinatí a hadi), ptáci rostliny ojediněle: Fragaria orientalis
Vznik pohlavních chromozomů: diferenciace některého autozomového páru primitivní pohlavní chromozomy homomorfní: nelze je rozlišit morfologicky, pouze studiem genů vázaných na pohlaví nebo vyvoláním tzv. pohlavního zvratu (syngamie je zde dosud labilní) Difererenciace pohlavních chromozomů má dvě důležité složky: 1. Suprese crossing overu mezi párovým a neprovým pohl. chromozomem. Rekombinace je nevýhodná pro geny, jejichž alely mají různý efekt u obou pohlaví. Proto preferována vazba těchto alel. To se může týkat nejen genů, kt. ovlivňují produkci gamet, primární nebo sekundární pohl. znaky, ale i genů kt. podmiňují normální fenotypové znaky Mechanismus suprese: mutace v genech, chromozomové aberace (např. inverze, translokace, duplikace) 2. Degenerace nepárového pohlavního chromozomu Mullerova rohatka (* Muller 1964, název pochází od Felsensteina *1974): model jednosměrné progresívní akumulace ztrátových mutací na nepárovém pohlavním chromozomu - v tomto modelu geny na Y(W) kompletně vázány. Náhodně z populace eliminován chromozom bez ztrátové mutace, pak s jednou ztrátovou mutací atd. Ztracená alela (úsek) nemůže být získána z pár. pohl. chromozomu vzhledem k supresi rekombinace mezi pár. a nepár. pohl. chromoz.
Premisy modelu: A. v populaci se vyskytuje Y(W) chromozom s 1, 2, 3. . . n ztrátovými mutacemi v různých lokusech B. selektivní výhody jednotlivých lokusů jsou poměrně malé C. velikost populace je dostatečně malá vzhledem k mutační rychlosti Degenerace nemůže být jen důsledkem prostého hromadění recesivních alel. Spolupůsobí další procesy: kumulace vysoce repetitivní DNA, transpozabilních elementů, hitchhiking. Další degenerace posilována také kompenzací funkčními protějšky na X popř. regionální supresí exprese na Y. Pohlavní chromozomy se rozrůzňují morfologicky (heteromorfie), v extrémním případě se Y(W) zcela ztrácí.
Vznik neopohlavních chromozomů: přestavbami mezi stávajícími pohlavními chromozomy a autozomy resp. rozpadem stávajících pohlavních chromozomů Následuje opět rozšíření restrikce rekombinací , degenerace nepárového pohlavního chromozomu atp. vznik: zejm. reciproké translokace, Robertsonovy translokace, rozpad chromozomů - významný efekt: fixace heterotického efektu u heterogametického pohlaví - významná reprodukční bariéra - možná reverze systému X 0 na XY - někdy vede ke značnému snížení počtu autozomů
Blaps polychresta (Tenebrionidae) Degeneraci nepárového pohlavního chromozomu doprovází inaktivace pohlavních chromozomů v meiotickém dělení heterogametického pohlaví. V profázi I leží pohlavní chromozomy na periferii jádra a jsou více spiralizovány než autozomové bivalenty (tzv. sex vezikul). Vyšší superspiralizace se manifestuje odlišnou barvitelností, tzv. heteropyknózou. Někdy jsou pohlavní chromozomy inaktivovány již v mitózách které předcházejí meiotickému dělení. Význam inaktivace v meiotickém dělení: - posiluje supresi rekombinace mezi párovým pohlavním chromozomem a alozomem - inaktivace transkripce
Neobvyklé systémy pohlavních chromozomů systém X 1 X 20 u pavouků, některých lasturnatek, hmyzu, hlístic - u organismů s holocentrickými chromozomy (ploštice, hlísti) většinou fragmentací, sekundární systém - u pavouků pravd. evolučně původní, vznikl zřejmě z modu X 0 nondisjunkcí a následným rozrůzněním obou X-ů dalšími nondisjunkcemi vznikají u pavouků systémy X 1 X 2 X 30, X 1 X 2 X 3 X 40, ty jsou však vzácnější (systém X 1 X 20 u cca 75% dosud studovaných druhů) Někteří sklípkani mnoho X, X 1 -X 130 sklípkan Macrothele gigas
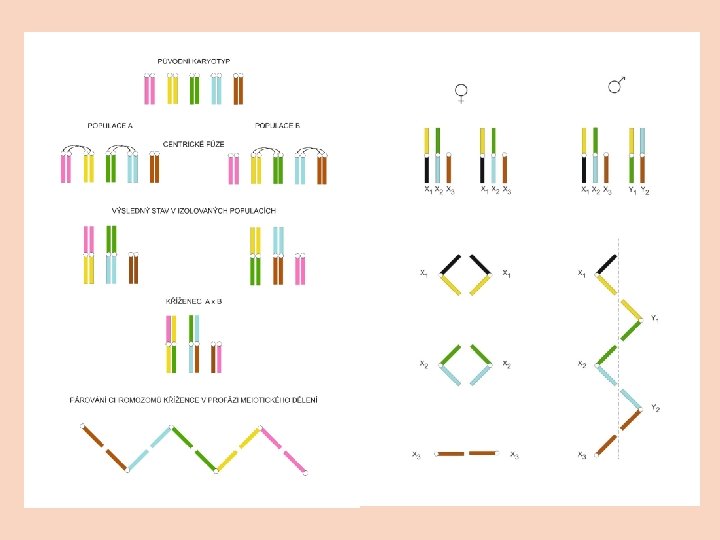
skákavky Habronattus systém X 1 X 2 Y sociální maloočka Delena cancerides - systém X 1 X 2 X 3 X 4 X 5 X 6 Y 1 Y 2 Y 3 Y 4 vznikl sérií fúzí
Monotremata Chromozomové řetězce (ptakopysk X 1 -5 Y 1 -5) Homologie s chromozomy ZW (u předků savců byl pravděpodobně systém Abraxas)
Monotremata Chromozomové řetězce (ptakopysk X 1 -5 Y 1 -5) Homologie s chromozomy ZW (u předků savců byl pravděpodobně systém Abraxas) Dva modely vzniku A centrické fůze a hybridizace (podobně jako u Delena cancerides) B sekvence translokačních událostí
Šťovík Rumex acetosa - XY 1 Y 2 z původního XY studium párování pomocí EM: Y 1 a Y 2 isochromozomy původního metacentrického Y primitivní žába Leiopelma hochstetteri Y 0/00 - Y vznikl stabilizací B chromozomu někteří pěvci (zebřičky) - univalent který se dědí jen v samčí linii (buď stabilizovaný B chromozom nebo výsledek soupeření původního typu Abraxas a nově vznikajícího systému Drosophila, který se však nakonec neuplatnil. Někdy existuje v populaci více konkurenčních heterogametií, většinou u jednoho pohlaví. Druhé pohlaví je pak jednotně homogametické. V rámci druhu můžeme najít i populace které se mohou lišit i typem heterogametie s ohledem na pohlaví. Jedná se o tzv. multiple factor systems. samice samec aabb Aabb aa Bb Výskyt: moucha domácí, pakomáři, mečovky, lumíci Tyto situace dokládají soupeření starého a nově vznikajícího chromozomového určení pohlaví. Pokud jeden systém nabízí vyšší fitness, následuje disruptivní selekce a jeden ze systémů je z populace vytlačen
Kompenzace dávky na pohlavních chromozomech další jev, který doprovází degeneraci nepárového pohlavního chromozomu - Drosophila: chromozom X samce 2 x aktivnější (s výj. některých lokusů jako NORy - jsou na X i Y, žloutkových genů - jen u samice a genů, které jsou pravděpodobně autozomového původu - tato regulace zůstane zachována i po přenesení fragmentů X na autozom - savci: lyonizace (inaktivace jednoho X u samičího pohlaví, embryonální vývoj) - omezena na somatické buňky (* Lyonová 1962) - inaktivovaný X leží pod cytoplazmatickou membránou, fakultativně heterochromatinický (Barrovo tělísko, * Barr a Bertram 1949). - inaktivovaný X se replikuje později - vačnatci - inaktivován paternální X, eutheria - inaktivace náhodná u některých bodlínů, myší je inaktivovaný X ztracen (somatické tkáně X 0) totéž v některých buněčných populacích starších žen (lymfocyty)
segmentované neutrofily - inaktivovaný X tvoří výběžek jádra - drumstick výjimky z náhodné inaktivace: - některé tkáně - pokud jeden X aberovaný, je inaktivován přednostně - pokud jeden X translokován na autozom: inaktivován zdravý X - u mezidruhových kříženců inaktivován vždy ten X který se replikuje později
Některé části X (zejména pseudoautozomová oblast) nejsou inaktivovány. inaktivace je řízena z tzv. inaktivačního centra (v proximální části Xq). Rozhodující regulační roli v tomto procesu hraje RNA genu XIST. Tento gen je aktivní na inaktivovaném X. Jediná molekula RNA genu XIST dokáže inaktivovat až 2 Mb na X. Molekulární mechanismus inaktivace: methylace cytozinových zbytků určitých sekvencí inaktivovaného X, methylace N konce histonu H 3, hypoacetylace histonu H 4 Další příklady somatické inaktivace pohlavních chromozomů: samice motýlů (ZW), inaktivován je chromozom W
Párování heteromorfních pohlavních chromozomů v meióze: - prostřednictvím chiazmat v homologickém úseku (člověk, PAR pseudo autosomal region) Pokud není dostatečná homologie, vyvíjejí se sekundární zůsoby párování: 1. Značně pozměměný synaptonemální komplex (někteří pavouci) 2. heterochromatinové bloky X a drobný Y u brouků - párující figura připomíná parašutistu s padákem 3. tzv. segregační tělíska - zvláštní materiál, odlišný od SK (X a Y Drosophila) 4. trakci pohlavních chromozomů zabezpečují autozomy (škrkavky) 5. mezi pohlavními chromozomy se vyvíjí separátní svazek mikrotubulů, který zabezpečuje rozchod krtonožka Neocurtilla hexadactyla - systém X 1 X 2 Y, svazek mezi X 1 a Y
Haplodipoidie - zvláštní způsob determinace pohlaví, zahrnuje partenogenetický vznik samců (arrhenotokie) z neoplozených vajíček - samice 2 n, samec n: samici můžeme formálně považovat za XX, samce za X 0 - vznikla u různých živočišných skupin: vířníci, blanokřídlí, třásněnky, někteří stejnokřídlí (molice, červci), primitivní brouk Micromalthus debilis, někteří roztoči Důsledky haplodiploidie: 1. menší heterozygotnost - škodlivé alely se odstraní u samců 2. poměr pohlaví nemusí být 1: 1 3. zvyšuje se příbuznost mezi samičími členy populace (preadaptace pro socialitu) 4. kompenzace dávky pomocí polyploidizace některých tkání u samců Evoluce haplodiploidie zahrnuje inaktivaci samčí sádky, případně její odvržení v somatických tkáních samců (parahaploidie). Tyto mezistupně nejlépe zachyceny u červců a roztočů čel. Phytoseiidae.
Haplodiploidie - predispozice k evoluci eusociality u blanokřídlého hmyzu 1. Vychílení poměru pohlaví 2. Nárůst inkluzivní fitness (Teorie sobeckého genu)
Thelytokie - samci úplně odbouráni, partenogeneticky vznikají jen samice - odbourání sexuálního rozmnožování, vznikají klony, které se dále rozrůzňují různými mutacemi Význam: - fixace heterozygotnosti nebo naopak postupně kompletní homozygotnost - fixace polyploidie - přizpůsobeni se neměnícím podmínkám, na okrajích areálu v extrémních podmínkách pro druh (zvyšuje možnosti disperze při nepatrné populační hustotě) - thelytokní taxony mnohdy (ale ne vždy) slepou uličkou evoluce Typy thelytokie: Automiktická - zachována meióza. Postup k homozygotnosti, který různě rychlý podle toho které produkty meiotického dělení splývají Apomiktická - meióza odbourána, vajíčka vznikají mitoticky. Fixuje heterozygotnost organismu která je dále posilována novými mutacemi
Genomový imprinting - určitá část genomu (chromozomy, jejich úseky či jednotlivé geny) fungují odlišně v závislosti na rodičovském původu Mikromanipulační konstrukce aberantních zygot u myší: A. se dvěma samičími prvojádry: abnormality obalů, embryo normální (u lidí ovariální teratom) B. se dvěma samčími prvojádry: obaly relativně normální, embryo abnormální (u lidí hydatiformní mola) Význam: u savců ve vývoji placentace (diference ve funkci otcovského a mateřského genomu v embryu a placentě - zajištění tolerance). Otcovský a mateřský organismus má tedy odlišnou úlohu při vývoji embrya obalů a placenty. Existenci imprintingu potvrzuje i uniparentální disomie různých chromoz. párů (např. oplozením disomické a nulisomické gamety, tzv. gametická komplementace). Imprinting se rovněž manifestuje u jedinců s delecemi, v závislosti na tom, od kterého rodiče delece pochází (Prader-Willi - delece na otcovském, Angelmannův syndrom - na mateřském chromozomu, jinak jde o stejnou deleci - odlišné fenotypy) Vznik: gametogeneze (změna methylačního patternu sekvencí)