Juhuslikud sndmused populatsioonigeneetikas Binoomjaotus Viljastuvad gameedid juhuslik vike

Juhuslikud sündmused populatsioonigeneetikas

Binoomjaotus Viljastuvad gameedid – juhuslik väike valim suurest hulgast gameetidest Kui tegemist AA homosügoodiga, siis kõik gameedid A tüüpi Kui tegemist Aa heterosügoodiga, siis oodatav 0. 5 A ja 0. 5 a aga Binoomsed tõenäosused saada A alleel (1/2 tõenäosuse juures) 10 -se valimi juures

Binoomjaotus N=10 Kui visata münti n korda siis mis on “kulli” ja “kirja” tõenäosused: N=100 97, 8% 96, 6% Tõenäoline Oodatav 95%, 99%. . Ebatõenäoline kokkulepe
 jaotus konveier")
Binoomjaotus Mittejuhuslik (predetermineeritud) jaotus konveier

Juhuslik geneetiline triiv Ühesuguse kohasusega genotüüpide sagedus muutub ajas juhusliku geneetilise triivi tõttu genotüüp AA Aa aa kohasus 1 1 1

sim Juhuslik geneetiline triiv Geenitriivi simulaatorid internetis http: //darwin. eeb. uconn. edu/simulations/drift. html 10 populatsiooni N= 1000 Üle suure hulga populatsioonide jääb p samaks (kui ei mõjuta kohasust). Suureneb sageduste dispersioon. Igas põlvkonnas: var(p)=pq/2 N See on Wright – Fisher mudel.

Juhuslik geneetiline triiv Kromosomaalne inversioon “PP” erinevus suurte ja väikeste Drosophila populatsioonide vahel 10 väikest (n=20 18 kuud ja 10 suurt n=4000) 18 kuud Dobzhansky ja Pavlovsky väikene populatsioon kaldub suurema tõenäosusega eemale oodatud keskmisest
: Uue populatsiooni rajamine väikese arvu (äärmisel juhul")
Juhuslik geneetiline triiv: rajajaefekt E. Mayr (1963): Uue populatsiooni rajamine väikese arvu (äärmisel juhul üheainsa viljastatud emase) isendite poolt, kes kannavad endas vaid väikest osa vanempopulatsiooni geneetilisest varieeruvusest. rajaja “efekt” rajaja “sündmus” rajajaefekt (founder effect)
 seni asustamata piirkondade koloniseerimisel nt. pärinevad pea kõik Tristan")
Juhuslik geneetiline triiv: rajajaefekt A) seni asustamata piirkondade koloniseerimisel nt. pärinevad pea kõik Tristan da Cunha saare elanikud (ca 300) ühest Šoti perekonnast (kapral William Glass), kes saabus saarele 1817, + mõningad hilisemad migrandid
 B) pudelikael: üksikud indiviidid jäävad ellu ja taasloovad populatsiooni")
Juhuslik geneetiline triiv: pudelikael (bottleneck) B) pudelikael: üksikud indiviidid jäävad ellu ja taasloovad populatsiooni (ekspandeeruvad)

Right Whales Eubalaena glacialis, "good, or true, whale of the ice. " Lõunavaal Eubalaena spp. About 400 North Atlantic right whales exist today.
 ja näljad. Eesti rahvaarv kahanes 365000 -> ca.")
Kui oluline pudelikael? Põhjasõda (1700– 1721) ja näljad. Eesti rahvaarv kahanes 365000 -> ca. 130 000
 sõltub rajajate arvust ja")
Juhuslik geneetiline triiv: rajajaefekt/pudelikael Populatsiooni geneetiline mitmekesisus (heterosügootsus / homosügootsus) sõltub rajajate arvust ja alleelisagedustest vanempopulatsioonis Diploidne 2 -alleelne lookus Homosügootne tõenäosus = (p 2)N + (q 2)N
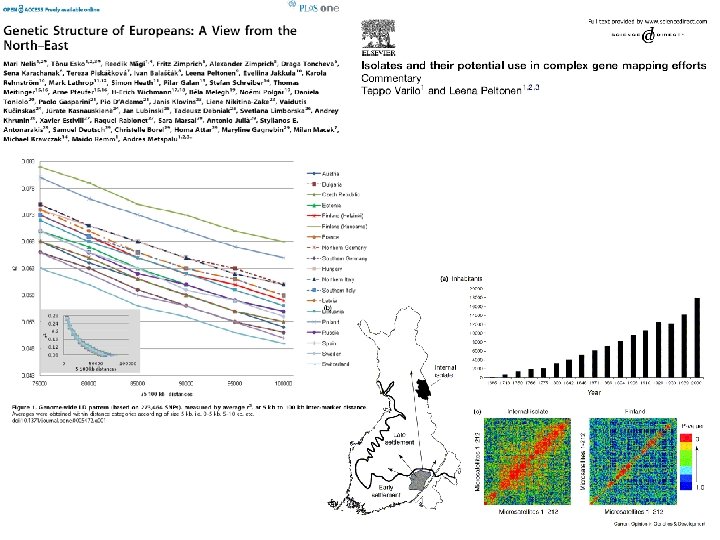

Juhuslik geneetiline triiv: rajajaefekt/pudelikael Tõenäosus monomorfsuse tekkeks väike, kui just tõesti ainult üks või paar rajajat ei juhtu olema. Isegi “käputäie” rajajate puhul on suur tõenäosus, et säilub “enamus” vanempopulatsiooni alleelidest (2 -3 alleelsetes lookustes), ent oluline efekt on nende alleelide sageduste muutumisele (stohhastika). Oluliselt suurem efekt multialleelsete lookuste korral (nt. mitokondri haplotüüpide rikkus), kus rajajate arv otseselt piirab erineva tüübiga järglaste edasikandumist, nt. kui algpopulatsioonis on sadu erinevaid haplotüüpe, siis 5 -10 rajaja korral säilub kindlasti vähem kui 10% geneetilisest mitmekesisusest, samas bialleelsete lookuste juures on kadu kvaliteedis vähemärgatav.
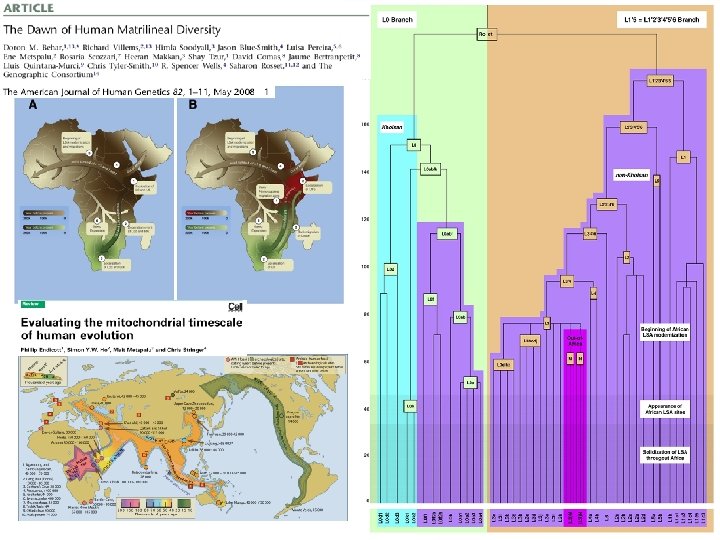

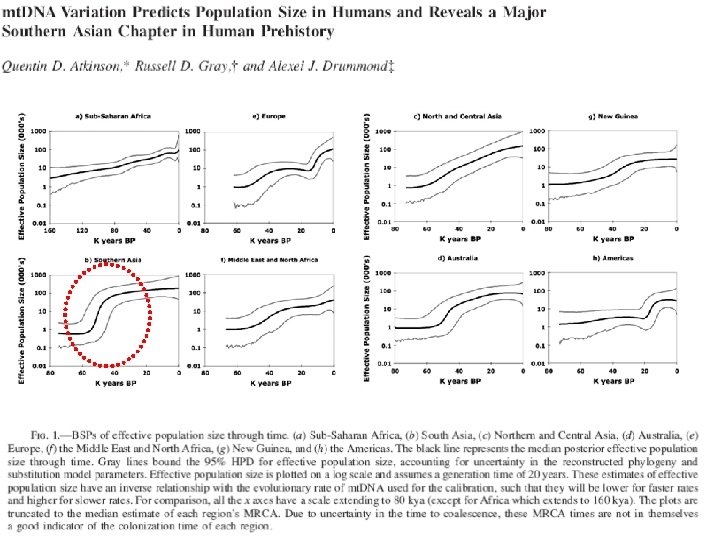
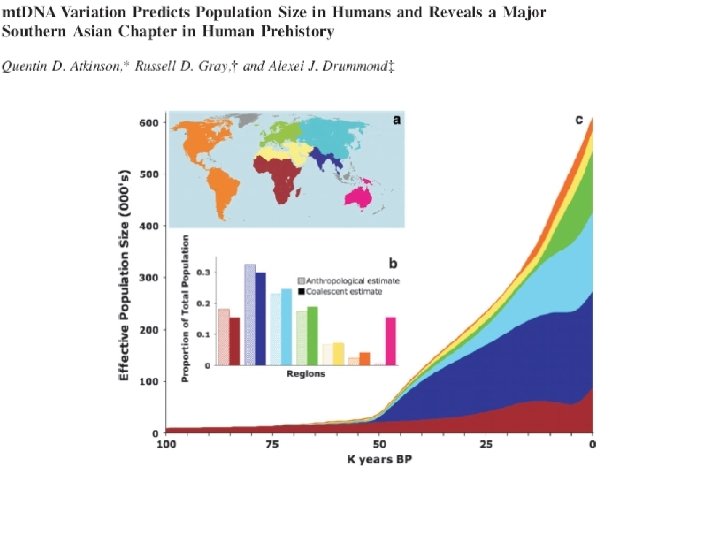
Juhuslik geneetiline triiv: rajajaefekt/pudelikael ~40 -65 KYA
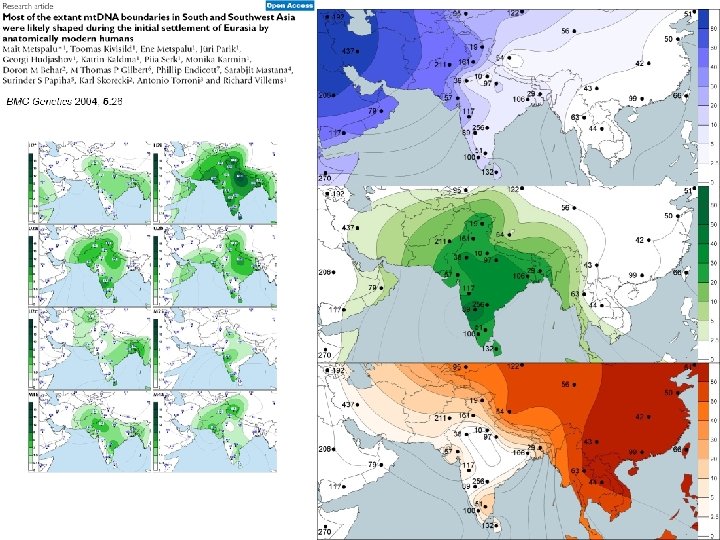
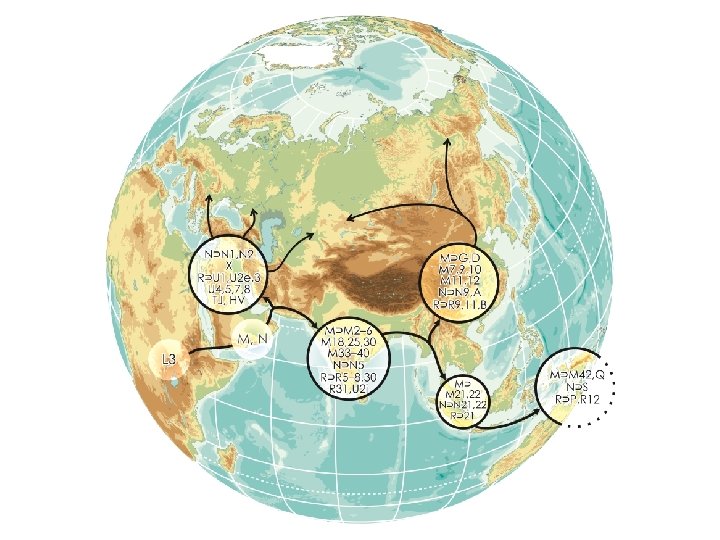

Juhuslik geneetiline triiv: rajajaefekt/pudelikael ~40 -65 KYA

Peopling of the Americas: a mt. DNA based reconstruction E. Tamm et al. 2007 PLo. S ONE epub 5/09

Juhuslik geneetiline triiv: rajajaefekt/pudelikael Both the extent and location of the maternal ancestral deme from which the Ashkenazi Jewry arose remain obscure. Here, using complete sequences of the maternally inherited mitochondrial DNA (mt. DNA), we show that close to one-half of Ashkenazi Jews, estimated at 8, 000 people, can be traced back to only 4 women carrying distinct mt. DNAs that are virtually absent in other populations, with the important exception of low frequencies among non-Ashkenazi Jews. We conclude that four founding mt. DNAs, likely of Near Eastern ancestry, underwent major expansion(s) in Europe within the past millennium.

Juhuslik geneetiline triiv: rajajaefekt/pudelikael Paarialleelse lookuse puhul on rajajaefekti peamine tulemus alleelisageduste muutumine Nt. Lõuna Aafrika valgete populatsioon (Afrikaner) pärineb umbes laevatäiest hollandlasest, 1652. Praegu u. 2. 5 M elanikkonnast 1 M kannab 20 algrajaja perekonnanime. Ühel rajajatest oli haruldane Huntingtoni tõbi – praegu selles populatsioonis sagedane, keskeas letaalne dominantne haigus Või porphyria variegata, samuti dominantne autosoomne geen. Ei kannata barbituraate (seega polnud oluline enne moodsat meditsiini). Täna 30, 000 ja ka tagasi viidav Gerrit Jansz ja Ariaantje Jacobs-i saabumisele 1685 ja 1688.

Juhuslik geneetiline triiv: rajajaefekt/pudelikael Iseenesest rajajaefekt ei suurenda mingite haiguste esinemise tõenäosusi rajatud populatsioonis – paljude teiste haiguste tõenäosus võib vastupidi olla väga väikene, juhuslikult mõni aga võib saada teistega võrreldes suurema sageduse (tänu juhuslikule geneetilisele triivile)

Juhuslik geneetiline triiv: rajajaefekt/pudelikael Finland's demographic history has led to a unique catalogue of genetic diseases. More than 35, mostly recessive diseases, are highly enriched in Finland. Other diseases, such as phenylketonuria and cystic fibrosis are almost non-existing.

Juhuslik geneetiline triiv: rajajaefekt/pudelikael

Juhuslik geneetiline triiv: neutraalse evolutsiooni kiirus 6 alleeli 3 indiviidi iga üksiku alleeli fikseerumise tõenäosus on väga madal, diploidsetel 1/(2 N) ehk siis siin 1/6 Kaotsiminemise tõenäosus suur (2 N-1)/2 N ehk siis siin 5/6

Juhuslik geneetiline triiv: neutraalse evolutsiooni kiirus Fikseerumise tõenäosus, samuti igale uuele mutatsioonile on 1/(2 N), Neutraalsed mutatsioonid tekivad kiirusega u (jälle, mutatsioonikiirus geeni kohta), Ühes põlvkonnas (kus on 2 N alleeli) tekib 2 Nu uut mutatsiooni Evolutsioonikiirus seega korrutis 2 Nu ja nende fikseerumise tõenäosusest 1/(2 N) kus populatsiooni suurus välja taandub = u -> neutraalne evolutsioonikiirus võrdub mutatsioonikiirusega, rippumata populatsiooni suurusest

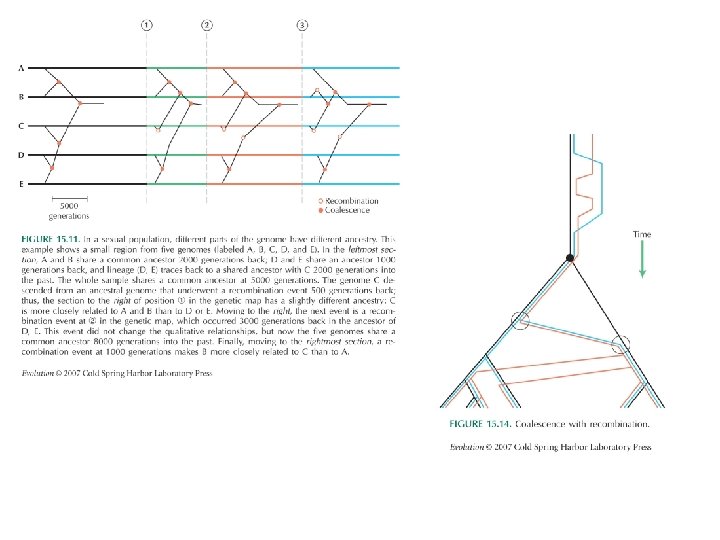
Juhuslik geneetiline triiv: coalescence: koalestents, koalestseerumine Vaadates triivi ajas tagasi näeme tänaste alleelide koalestseerumist eellasalleeli Mida väiksem on populatsioon, seda kiiremini ajas tagasi minnes koalestseerumispunkti jõuame. …sest seda suurem on iga alleeli tõenäosus igas põlvkonnas nulli triivida

Juhuslik geneetiline triiv: coalescence: koalestents, koalestseerumine konstantne Ne Suurem osa koalestseerumisi toimub kiiresti. Keskmiselt on 2 Ne põlvkonna järel alles 2 eellast ja läheb veel 2 Ne põlvkonda enne kui need koalestseeruvad. pudelikael Pudelikael võimendab koalestseerumiste koondumist. Sisuliselt tegemist väiksema Ne ga Sest tagasi minnes on igas põlvkonnas N sama ja valdav enamus koalestseerumisi toimub tänaseks kadunud geenide vahel. See kõik kehtib keskmiselt, tegelikkuses on varieeruvus on suur kasvav Ne Kasvavas populatsioonis on geenide kaotsimineku tõenäosus väiksem, mistap on ka koalestseerumised suhteliselt sügavamal Aeg põlvkondades mutatsioonid
")
Inferring past effective population sizes in 8 world regions using Bayesian skyline plot (BSP) Internally calibrated clock (hg Q in Sahul at 45 KYA) The growth curves are derived from changes in the inferred lineage coalescent rate through time.

Juhuslik geneetiline triiv: coalescence: koalestents, koalestseerumine Tavaliselt me ei saa genealoogiat live’is jälgida vaid taastame selle säilunud geenivariantidest. Seega näeme vaid nende eellasvariante ja mitte kogu varieeruvust igas põlvkonnas. Siin (b) on erand, kus HIV (a) HIV gag geeni puu 200 env geeni genealoogiat patsiendilt jälgiti ühel patsiendil 7 aasta jooksul

Juhuslik geneetiline triiv: HW ja triiv HW tasakaal ei ole tasakaaluline “väikestes” populatsioonides suhteline mõiste, võrreldes gameetide arvuga on kõik populatsioonid väikesed eeldus: alleelisageduste kõikumine mingi keskmise ümber sagedus tegelikkuses: alleelisagedused 1 kipuvad fikseeruma. Kui fikseerumine on toimunud siis tagasiteed ei ole. Siis on tõeline stabiilne (0/1) tasakaal. 0 aeg

Juhuslik geneetiline triiv: HW ja triiv Tegelikult ka suures populatsioonis, iga põlvkond määratakse HW tasakaaluolek uuesti ja see määrab vaid oodatava, mitte tingimata realiseeruva sageduse vastavatele alleeli ja genotüübi sagedustele sagedus 1 0 aeg Lõpmata suures ideaalpopulatsioonis need muutused lõpmata väikesed ja teoreetiliselt siis ei kehti, sama kui vaadelda suhteliselt suuri populatsioone suhteliselt väikeses ajaskaalas

Juhuslik geneetiline triiv: Marss homosügootsusele Homosügootsus / Heterosügootsus kuidas hinnata? juhuslik geneetiline triiv viib neutraalsed lookused varem või hiljem homosügootsesse seisundisse. Ja teeb seda kiiremini väiksemates populatsioonides.
 = 1 – f (fikseerunud alleelide,")
Juhuslik geneetiline triiv: hetrosügootsus /gene diversity H (heterosügootsus) = 1 – f (fikseerunud alleelide, e. homosügootide osakaal populatsioonis) f = A 1 A 1 + A 2 A 2 +. . . An, kokku 2 N alleeli -> gameeti ->. . p 2 q 2 e. H = 1 – (p 2 + q 2 +. . ) = 1 – pi 2 HW tasakaalu korral on H sama mis arvuliselt heterosügootide osakaal (nt. Aa) Aga nt: 50 AA, 0 Aa, 50 aa H = 0. 5 Sest H on olemuselt tõenäosus, et populatsioonist juhuslikult (ühest lookusest) valitud kaks geeni on erinevad alleelid.
: Üks populatsiooni sisese")
Juhuslik geneetiline triiv: hetrosügootsus /gene diversity Gene diversity (expected heterozygosity/oodatud heterosügootsus): Üks populatsiooni sisese geneetilise mitmekesisuse mõõte: tõenäosus, et populatsioonist juhuslikult valitud kaks geeni (ühest lookusest) on erinevad alleelid. Tavaliselt HE, või Hexp, algataja Bruce Weir. Üks lookus, k alleeli m lookust, k alleeli

Juhuslik geneetiline triiv: hetrosügootsus /gene diversity Populatsioonisisesed genotüübisagedused p. S=0. 64; p 2=0. 41 q. F=0. 27; q 2=0. 08 r. I=0. 09; r 2=0. 01 f=0. 4247+0. 0843+0. 0151=0. 5241(homosügootide sagedus) H=0. 4759 (vaadeldud heterosügootide sagedus) H=1 -Sum(pi 2)=0. 50 (oodatud heterosügootide arv)
 statistiku F Antud")
Juhuslik geneetiline triiv: F statistikud Antud juhul võime kirjutada välja (üldise) statistiku F Antud näite korral, F= 0. 5 0. 48 =0. 004 Mida see arv väljendab?

Juhuslik geneetiline triiv: F statistikud F-statistikud: geneetilise struktuuri mõõdud, arendatud eelkõige Sewall Wright’i (1969, 1978) poolt. Sarnaselt statistical analysis of variance (ANOVA) FIS (Ka FS, inbriidingu koefitsient) populatsiooni geneetilise varieeruvuse osa, mis väljendub indiviiditi. Kõrged väärtused näitavad suurt inbriidingu osakaalu (palju homosügoote). Negatiivsete väärtuste puhul (outbriiding) palju heterosügoote (võrreldes HW oodatuga). -1. . . 1
 statistics HI – vaadeldud heterosügootide")
Juhuslik geneetiline triiv: F statistikud Weir and Cochram (1984) statistics HI – vaadeldud heterosügootide arv HS –populatsioonis oodatud heterosügootide arv (gene diversity) Nt 50 AA, 0 Aa, 50 aa; FIS=+1 0 AA, 100 Aa, 0 aa; FIS=-1 FIS = (HS-HI)/HS Mitme alampopulatsiooni korral rehkenda HS igas alampopulatsioonis ning võta nende kaalutud keskmine (alampopulatsioonide suuruste järgi) HI = HI 1*N 1 + HI 2*N 2 + HI 3*N 3 N

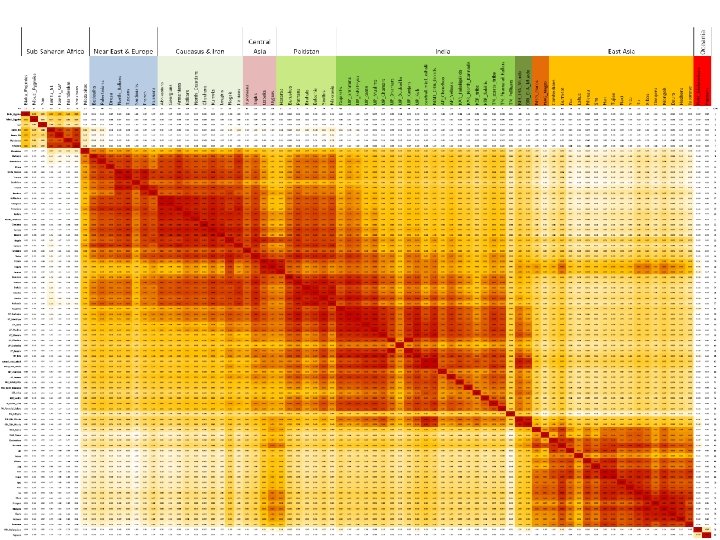
Juhuslik geneetiline triiv: F statistikud FST is the proportion of the total genetic variance contained in a subpopulation (the S subscript) relative to the total genetic variance (the T subscript). Values can range from 0 to 1. High FST implies a considerable degree of differentiation among populations. FST = (HT-HS)/HT Kus HS on alampopulatsioonide kaalutud keskmine heterosügootsus ja HT on populatsiooni üldine oodatud heterosügootsus Related measures: (theta) of Weir and Cockerham (1984) and GST of Nei (1973, 1978).
/HT=0.")
Juhuslik geneetiline triiv: F statistikud: Fst Platsentaarse aluselise fosfataasi näide =HT FST = (HT-HS)/HT=0. 06 Interpretatsioon: 6% populatsioonide vaheline 94% populatsioonide sisene varieeruvus. HS=0. 4134

Juhuslik geneetiline triiv: F statistikud: Fst


Juhuslik geneetiline triiv: F statistikud Kuigi FST kasutatakse eelkõige populatsioonide eristumise mõõtmisel, oli tema algne kasutusele võtmise eesmärk (Wright 1921) jälgida protsessi, mille läbi geneetiline mitmekesisus väheneb ja alleelid fikseeruvad (seetõttu ka F-statistika)

Juhuslik geneetiline triiv: mutatsiooni ja triivi tasakaal Mutatsioonid tagavad selle, et populatsioonid ei muutuks totaalselt homosügootseks vastavalt populatsiooni suurusele erinevad heterosügootsuse tasakaaluolekud (kuna (neutraalne) mutatsioonikiirus u on üldine) H* = 4 Nu + 1 H Seega, mida suurem populatsioon, seda suurem on heterosügootsus ehk seda suurem on mitmekesisus TEOREETILINE! Järgmises loengus vaatame kas ka tegelikult Nu

Juhuslik geneetiline triiv: Efektiivne populatsioonisuurus - N e Oluline on efektiivne populatsioonisuurus Ne mitte absoluutarv (loendus) N Populatsioonigeneetiliselt oluline see osa populatsioonist, mis määrab alleelide edasikandumise järgmisesse põlvkonda (Wright 1931) Populatsiooni efektiivne suurus on ideaalpopulatsiooni suurus, mis käitub juhusliku geneetilise triivi tingimusis sarnaselt meie uuritava populatsiooniga. Saab formaalselt arvutada • läbi alleelisageduste dispersiooni muutumise põlvkondade vahel • läbi inbriidingu koefitsiendi muutumise põlvkondade vahel The idealized case refers to a diploid population of N hermaphrodictic, selfcompatible, random mating adults with discrete generations and no age structure.

Juhuslik geneetiline triiv: Efektiivne populatsioonisuurus - N e sõltub, mitmetest asjaoludest: 1. Ebavõrdne sooline jaotus, Sewall Wright (1932): 4 Nm x Nf Ne = Nm + Nf 2. Populatsiooni suuruse kõikumised (pudelikael) 3. Väikesed paardumisgrupid (alamstruktuur), mis segavad panmiksist 4. Kattuvad põlvkonnad, vanuseline struktuur jne

Juhuslik geneetiline triiv: Efektiivne populatsioonisuurus - N e ühe isase korral, näiteks, Ne = 2 N/(1+N/2)=ca 4 Kui emaseid on 1000 siis 3, 99 Kui emaseid on 100 siis 3, 9 Kui emaseid on 10 siis 3, 4 näide, 96 lehma ja 4 pulli:

Juhuslik geneetiline triiv: Efektiivne populatsioonisuurus - N e Ne ja varieeruvus reproduktsiooni efektiivsuses võrdne reproduktiivne efektiivsus ehk varieeruvus=0 Ne=5 ebavõrdne reproduktiivne efektiivsus ehk suur varieeruvus=1 Ne=1

Juhuslik geneetiline triiv: Efektiivne populatsioonisuurus - N e kõrge varieeruvus reproduktiivses edukuses tähendab väikest Ne-d

Juhuslik geneetiline triiv: Efektiivne populatsioonisuurus - N e N fluktuatsioonid ja harmooniline keskmine näide:

Juhuslik geneetiline triiv: Efektiivne populatsioonisuurus - N e efektiivse populatsiooni suhe tsensusesse mõnedel kaslastel inimesel ~1/3

Juhuslik geneetiline triiv: mitmekesisus ja Ne Mutatsioonid tagavad selle, et populatsioonid ei muutuks totaalselt homosügootseks vastavalt populatsiooni suurusele erinevad heterosügootsuse tasakaaluolekud (kuna mutatsioonikiirus u on üldine) H* = 4 Neu + 1 H Seega, mida suurem Ne, seda suurem on heterosügootsus ehk seda suurem on mitmekesisus N eu

Juhuslik geneetiline triiv: mitmekesisus ja Ne Seega, neutraalse evolutsiooni korral peaks väiksema N e ga populatsioonid olema ühetaolisemad. Sama peab kehtima ka erineva pärandumismehhanismiga kromosoomidele, milledel on erinev Ne Ne Autosoomid: N X kromosoom: ¾ N Y kromosoom: ¼ N π exp 100% 75% 25% π 0, 000765 0, 000469 0, 000151 61% 20% enamvähem… Etteruttavalt, asi ei ole nii lihtne. Aga sellest kolmapäeval

Kokkuvõte Geneetiline triiv – väikestes populatsioonides alleelisageduste muutumine Rajajaefekt Mutatsiooni ja triivi tasakaal Efektiivne populatsioonisuurus
- Slides: 61