Evolcija Evolcijas etapi Organisko vielu izveidoans Organisko vielu



, CH 4, NH 3, H 2 O un")

.")


















 — A team of Penn State researchers has developed")












; • Eikariotu šūnas")









. Tās savieno hloroplastus vienotā tīklā, transportē proteīnus,")
 ir DNS")






- Slides: 75
Evolūcija
Evolūcijas etapi • • Organisko vielu izveidošanās; Organisko vielu polimēru izveidošanās; Organisko vielu kompleksu izveidošanās; Protošūnu izveidošanās; Metabolisko ceļu izveidošanās; Dzīvu šūnu izveidošanās; Endosimbioze; Sekundārā endosimbioze.
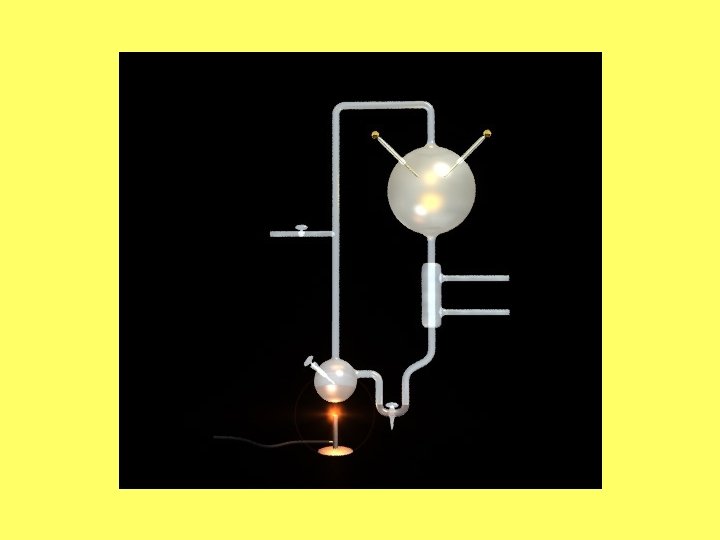
Organisko vielu izveidošanās • S. L. Millera rezultāti: • No: CH 4, NH 3, H 2 O un H 2 noslēgtā sistēmā izveidojas: urīnviela, aminoskābes (glicīns, alanīns u. c. ), pienskābe, etiķskābe un formaldehīds.
Organisko vielu izveidošanās No: HCN (cianīda), CH 4, NH 3, H 2 O un H 2 noslēgtā sistēmā izveidojas: urīnviela, aminoskābes (glicīns, alanīns u. c. ), pienskābe, etiķskābe, formaldehīds, cukuri, purīni, pirimidīni un porfirīni.
Organisko polimēru izveidošanās • Ūdens vidē parasti notiek nukleīnskābju, proteīnu, lipīdu un ogļhidrātu hidrolīze un sadalīšanās monomēros. • Paaugstināta temperatūra un ūdens iztvaikošana ļauj rasties polimēriem.
Organisko polimēru izveidošanās • Aminoskābes 1200 C – 2000 C veido garus polipeptīdus, (proteinoīdus). • Proteinoīdu veidošanos ūdens vidē stimulē arī cianīda un metālu klātbūtne.
Organisko polimēru izveidošanās • Nukleīnskābes veidojas no nukleotīdiem polifosfātu klātbūtnē. • Polifosfāti veidojas no fosfātiem 2000 – 3500 C.
Koacervācija • Polipeptīdi šķīdumā apvienojas pilienos d = 1 - 500 nm. • To nodrošina disulfīdu saites, hidrofobā mijiedarbība u. c. • Bieži izveidojas filmveida slānis. • Ja tas ir veidots no enzīmiem, tad var notikt enzimātiskas reakcijas. Oparins parādīja cietes veidošanos no cietes fosforilāzes, glikozes-1 -fosfāta klātbūtnē.
Proteinoīdu mikrosfēras • Fokss demontrēja, ka proteinoīdi ūdens vidē paaugstinātā temperatūrā veido mikrosfēras. • Tās saglabā vāju enzimātisko aktivitāti. • Elektronmikroskopiski pētījumi parāda, ka tās spēj pumpuroties.
Lipīdu dubltslānis • Fosfolipīdi un daži citi lipīdi ūdens vidē veido dubultslāni. Tas ir saistīts ar polāro ūdens molekulu iedarbību uz nepolārajām lipīdu molekulu daļām. • Bieži izveidotais dubultslānis veido nelielas lodveida struktūras. • Vezikulu virsmas kalpo vielu adsorbcijai. Citas vielas var uzkrāties vezikulu iekšienē. • Iespējama primitīva vezikulu reproducēšanas mehānisku spēku ietekmē (viļņi u. c. ). • Hipotētiskie apstākļi uz Zemes nebija piemēroti fosfolipīdu veidošanai. Tāpēc augsta fosfolipīdu koncentrācija, piemērota dubultslāņa veidošanai, ir apšaubāma.
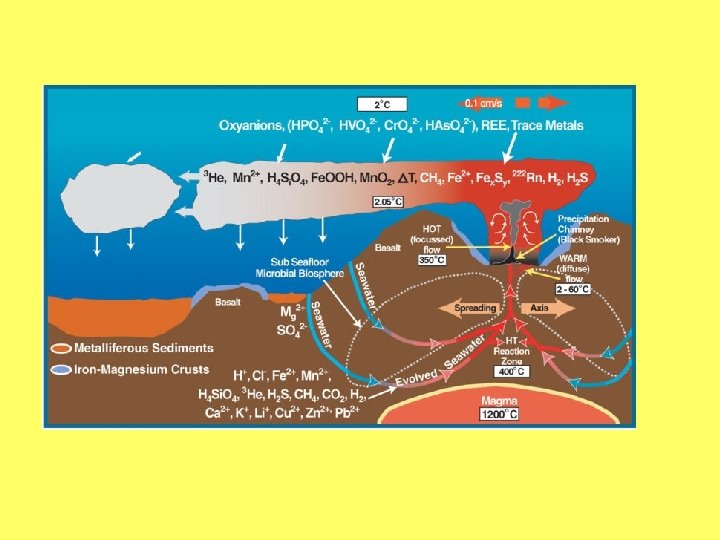
Sēravoti un dzīvības rašanās • Fe. S 2 • Organiskās vielas • Protošūna • Skurstenis un fosīlijas
Stromatolīts. Redzama karbonātus veidojoša zona J. Li, T. M. Kusky / Gondwana Research 12 (2007) 84– 100
Mikrobialīts no Ziemeļķīnas. Satur mikroorganismu atliekas. J. Li, T. M. Kusky / Gondwana Research 12 (2007) 84– 100
• Okeāna sēra avotu ieži ir poraini. • Tas ļauj metabolītiem uzkrāties un veidot plānu plēvīti, kas norobežo poru, tādējādi veicinot organisko vielu uzkrāšanos un protošūnu veidošanos. • Pēc šī modeļa no fosfolipīdiem veidotas membrānas būtu viens no pēdējiem posmiem protošūnu evolūcijā. • Tāpēc baktēriju, arhibaktēriju un eikariotu membrānām ir izveidojies atšķirīgs lipīdu sastāvs.
Mikrobialīts. e - Satur mikroorganismu sfēras; f –fosilas pavedienveida mikroorganismu atliekas dzelzs sulfīdā. J. Li, T. M. Kusky / Gondwana Research 12 (2007) 84– 100
Okeānos esošie sēra avoti satur sulfīdus. Šajos avotos ir augsta temperatūra un spiediens. Laboratorijās ir konstatēts, ka šādos apstākļos no CO 2 var veidoties etiķskābe, pirovīnogskābe un citi Krebsa ciklam raksturīgi savienojumi. Ir pierādīts, ka no pirovīnogskābes pakāpeniski var veidoties vairākas aminoskābes, piemēram, alanīns, serīns.
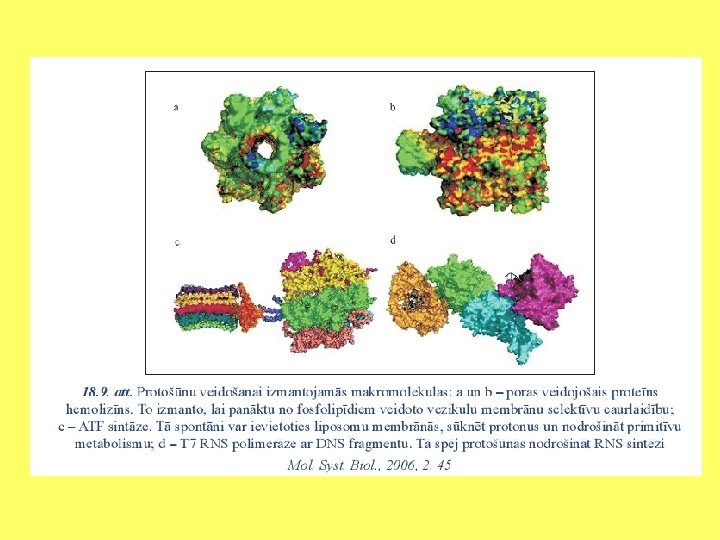
Protošūnas • Vezikulas varēja pārveidoties par protošūnām. • Tām jāvar: • transportēt jonus un organiskās vielas; • uztvert un transformēt enerģiju; • sintezēt molekulas. Lai to realizētu ir jābūt vezikulas apvalkam, kura centrālā daļa ir pildīta ar hidrofobām molekulu daļām un ārpuse ar hidrofīlām. Šajā slānī jābūt ievietotiem polipeptīdiem, lai tie kā enzīmi vai transporta olbaltumvielas palīdzētu veikt iepriekšminētās funkcijas. • Daudzi polipeptīdi spēj spontāni ievietoties membrānā un veidot kanālus.
Protošūnas • Ir pētīts polipeptīds, kas veidots no serīna un leicīna atkārtojumiem. Eksperimentālajā sistēmā bija oktāna slānis starp diviem ūdens slāņiem. • Peptīdu molekulas – pelēkas, ūdens : H – sarkans, O balts. Struktūra rāda, ka caur membrānu var pārvietot ūdens mlekulas.
Metaboliskie ceļi • Pirmās vielas, kuras varētu izmantot elektronu transportam protošūnās, bija porfirīni. • Veidojas Millera aparātā HCN klātbūtnē.
Metaboliskie ceļi http: //www. geocities. com/Research. Triangle/Node/5345/
Metaboliskie ceļi http: //www. geocities. com/Research. Triangle/Node/5345/
Metaboliskie ceļi • RNS molekulas glabāja pirmos gēnus un darbojās kā enzīmi. • Ribozīms – RNS molekula, kura darbojas kā enzīms. • Vienlaicīgi protošūnās atradās ribozīmi un polipeptīdi. • RNS varētu veidoties par DNS, ja protošūnā būtu enzīms reversā transkriptāze • Protošūna kura satur RNS vai DNS gēnus, veic replikāciju un proteīnu sintēzi ir saucama pa īstu šūnu. • Ribozīmus atklāja T. Cečs un S. Altmans un 1989 ieguva Nobela prēmiju ķīmijā.
Metaboliskie ceļi
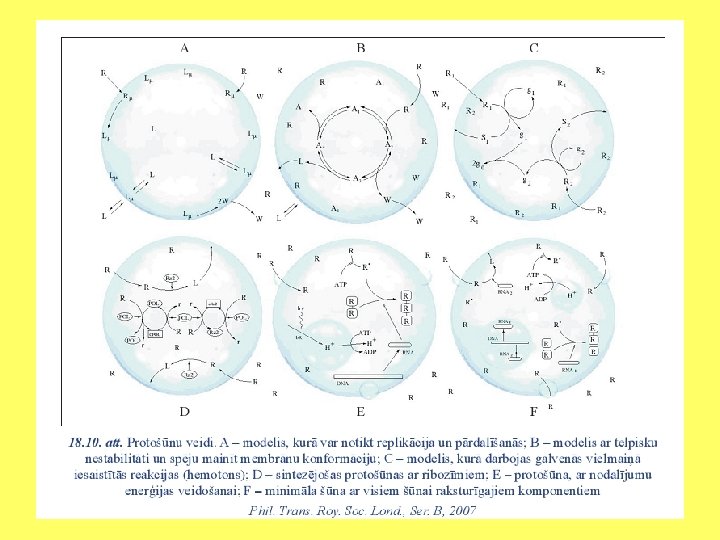
Mākslīgas protošūnas • Daudzās laboratorijās tiek veikti pētījumi, lai izveidotu mākslīgo šūnu. Tie norisinās divos virzienos: no augšas (top down) un no lejas (bottom up). • Pirmais veids paredz izpētīt vismazākos dzīvo šūnu genomus un to metaboliskās sistēmas. Otrajā gadījumā tiek pētīts, kādi elementi spēj pašsavākties, izveidot protošūnu, nodrošināt tajā noteiktas metaboliskās reakcijas un dalīšanos. • Pētījumos tiek veidoti šūnu uzbūves matemātiskie modeļi vai arī laboratorijas apstākļos izveidotas protošūnas no elementiem, kas spontāni var grupēties atbilstoši to ķīmiskajām un fizikālajām īpašībām.
Science. Daily (May 16, 2008) — A team of Penn State researchers has developed a simple artificial cell with which to investigate the organization and function of two of the most basic cell components: the cell membrane and the cytoplasm--the gelatinous fluid that surrounds the structures in living cells. The model cell developed in the lab of Christine D. Keating at Penn State uses as the cytoplasm a solution of two different polymers, PEG and dextran
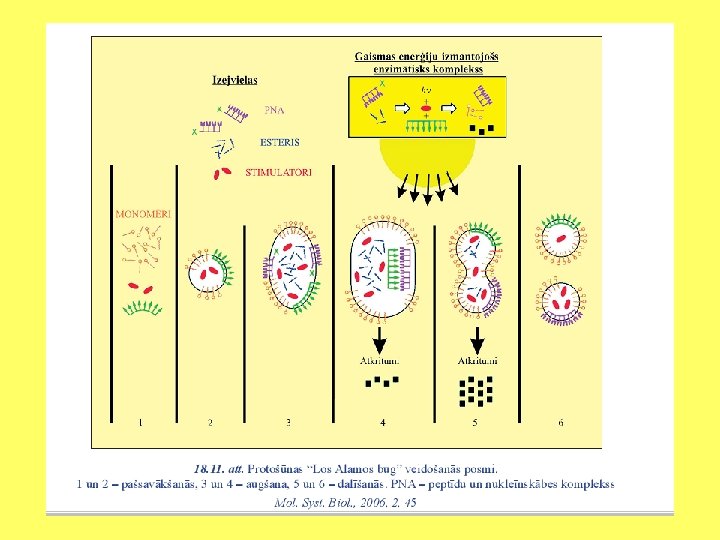
• Minimālās šūnas izveidošanas piemērs ir “Los Alamos bug”. Entonijs Forsters un Džordžs Čērčs ir aprēķinājuši, ka minimālais šūnas gēnu skaits ir 151. Protošunai “Los Alamos bug” ir 113 000 bāzu pāru, un tā satur 151 gēnu. • Pirmajā posmā sistēmā tiek ievadītas vielas, kas veido virsmu, katalizē reakcijas un darbojas kā genoms. • Otrajā posmā izveidojas micellas, kurās ievietojas katalizatori un uz kuru virsmas piestiprinās genoma funkcijas veicošie biopolimēri. • Trešajā posmā sistēmai pievieno izejvielas (virsmu veidojošās vielas, katalizatorus un nukleotīdu oligomērus). Esošā informācija ļauj nodrošināt protošūnas augšanu un replikāciju.
Protošūnas Radīšana Bioķīmiskā evolūcija Panspermija Mākslīgā šūna Protošūna un šūna Membrāna un koncentāciju atšķirība starp ārpusi un iekšpusi; Enerģijas ražošanas un pārveidošanas sistēma; Struktūrolbaltumu un enzīmu ražošanas sistēma; Informācija DNS un RNS veidā; Šūnu augšanas programma; Diferenciācijas programma; Informācijas apmaiņas programma; Nāves programma.
Protošūnu izcelšanās • Galvenā problēma ir tā, ka mūsdienu šūnām sastāvdaļas nerodas de novo. • Membrānas veidojas ET • Centriolas dalās Šūnas sastāvdaļu veidošanai vajadzīga matrice!!!
Protošūnu izcelšanās • Mūsdienu šūnas radušās no protošūnām. • Protošūnas radušās uz Zemes. • Protošūnas vai to daļas ir atnestas ar meteorītiem. • Kosmosā atrodamas dzīvas šūnas.
Protošūnas radušās uz Zemes 3, 8 miljardus gadu atpakaļ uz Zemes bija: • Gāzu maisījums atmosfērā un ūdenī. (H 2, N 2, CO 2, H 2 O) • Augstāka temperatūra kā mūsdienās • Augstāka radiācija kā mūsdienās. • Vairāk ultravioletās radiācijas kā mūsdienās.
• Viena no visvecākajām fosīlijām. • 3, 85 miljardi gadu veca. Atrasta Grenlandē. Prokariotu šūnas ir izmantojušas ar ūdeņradi bagātus savienojumus. NASA http: //www. solstation. com/life. htm
Protošūnas vai to daļas ir atnestas ar meteorītiem. • http: //popularmechanics. com/science/space/1999/7/alien_world 2/index 5. phtml
Galvenās dzīvo organismu grupas
Evolūcijas etapi • Pirmās prokariotu šūnas parādījušās 3, 8 miljd. g. atpakaļ. • Pirmās pierādāmās eikariotu šūnas parādījušās 1, 2 miljd. g. atpakaļ. • Senākās iespējamo eikariotu fosilās atliekas konstatētas 2, 1 miljd. g. vecos iežos. • Ievērojama skābekļa daudzuma palielināšanās atmosfērā notikusi 2, 1 miljd. g. atpakaļ.
Eikarioti Mičigana, Gripania, ~2. 1 miljardi gadu atpakaļ, šūna ar kodolu earth. usc. edu/~geol 150/evolution/ precambrianfossils. html
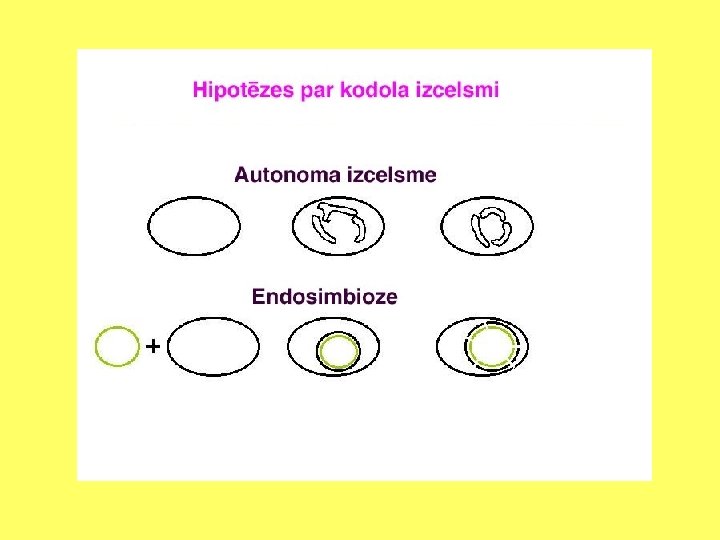
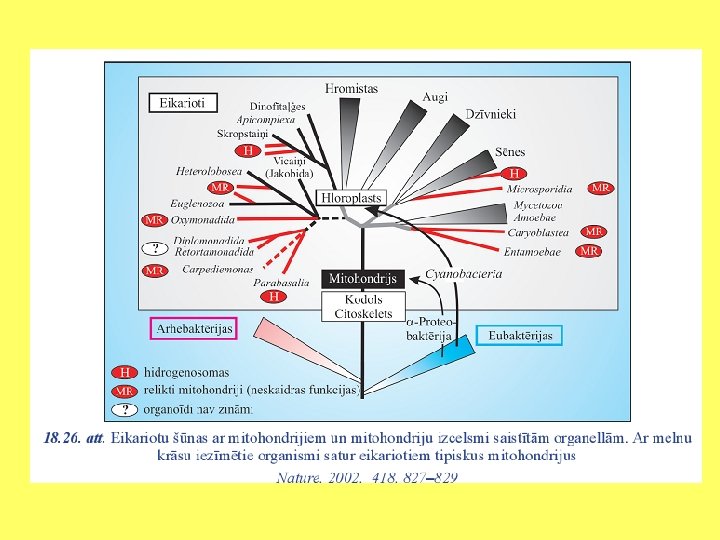
Endosimbioze un eikariotu evolūcija Plastīdas: Mereschkowsky 1910 Mitohondriji: Altman 1890, Wallin 1927 Autogēnā, pašsavākšanās hipotēze: Wilson 1925 Linna Margulis
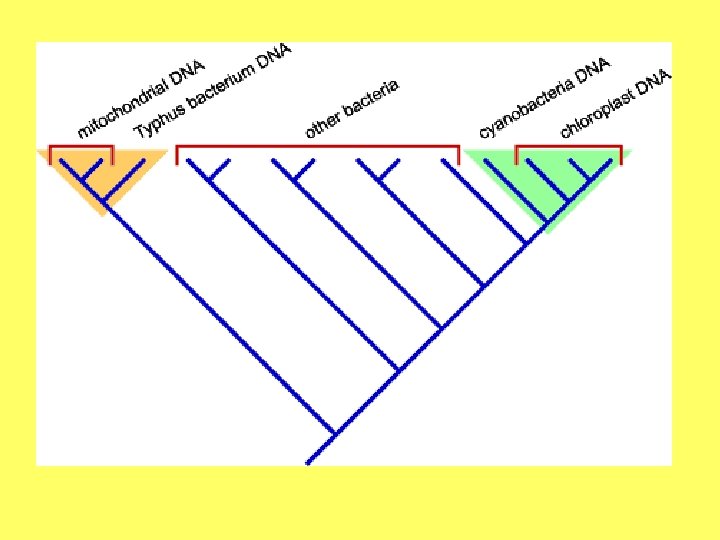
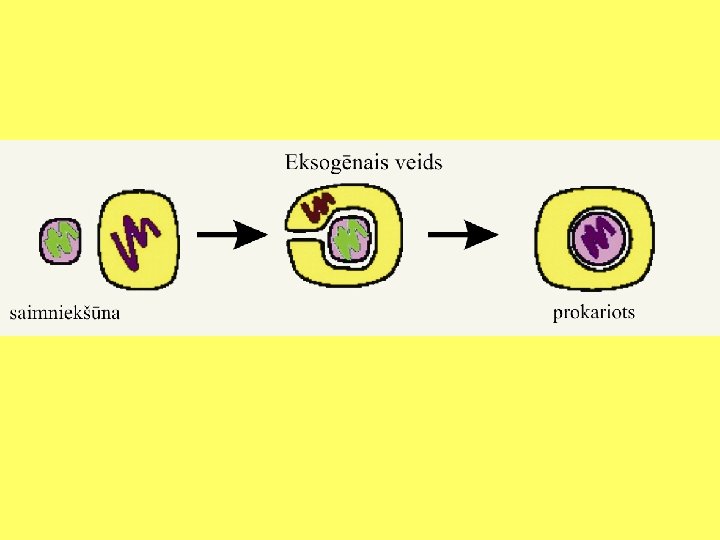
Endosimbiozes teorija • Eikariotu šūnu rašanos izskaidro endosimbiozes teorija (L. Margulis); • Eikariotu šūnas ir endosimbiotisks prokariotu šūnu komplekss. • Eikariotu šūnu priekšteči ir iekļāvuši šūnu iekšienē prokariotiskus simbiontus: • Aerobas baktērijas; • Fotosintezējošas baktērijas.
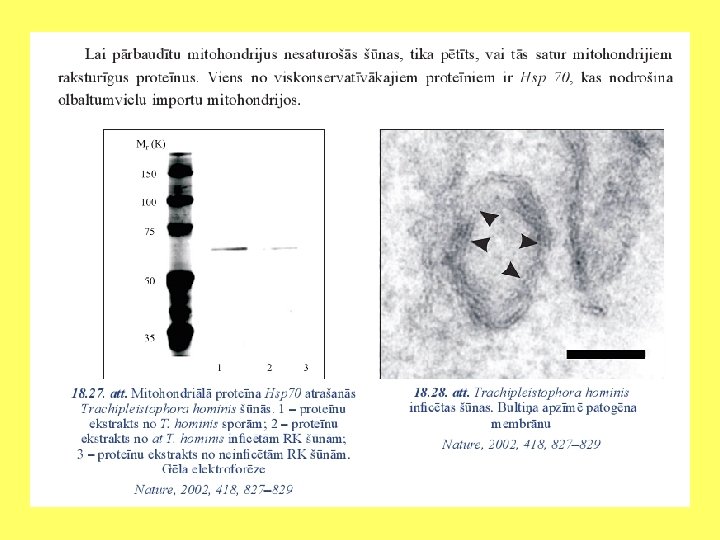
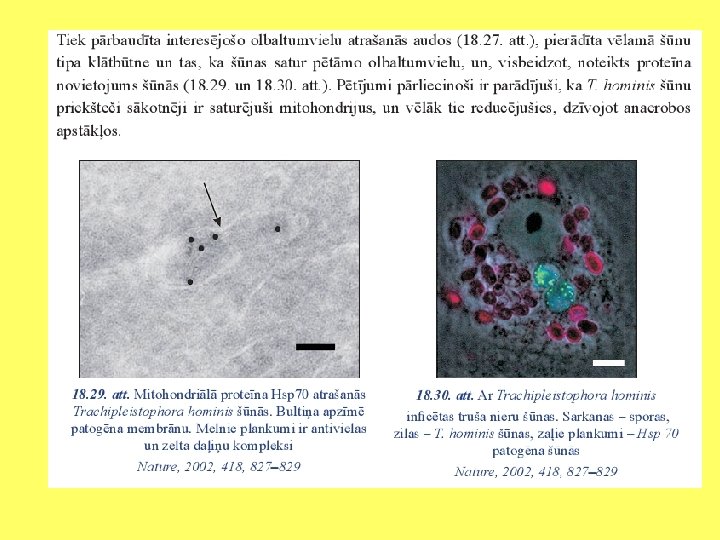
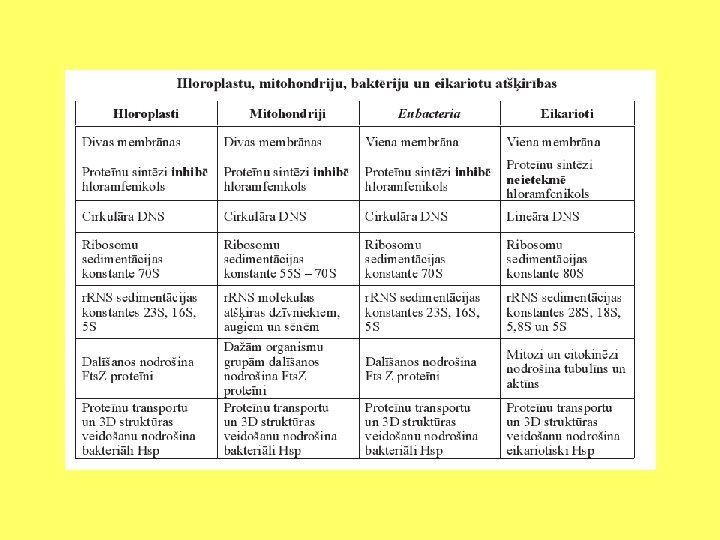
Endosimbiozes pierādījumi Divas membrānas Plastīdu un mitohondriju dalīšanās līdzīga kā prokariotiem Prokariotiska DNS Prokariotiska transkripcija un replikācija Prokariotiskas ribosomas Prokariotiskas siltuma šoka olbaltumvielas Līdzīgi kā baktērijas reaģē uz antibiotikām
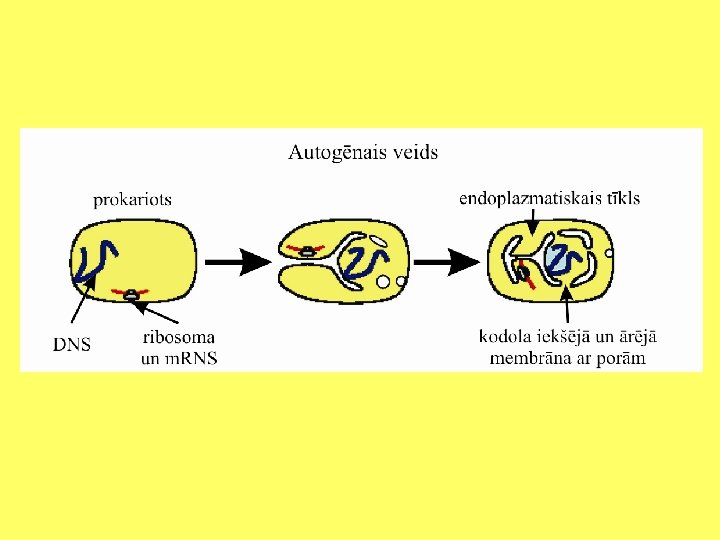
Svarīgākās eikariotu šūnu sastāvdaļas • Iekšējo membrānu sistēma: kodols, endoplazmatiskais tīkls, Goldži komplekss. • Lizosomas (Vakuolas). • Peroksisomas. • Plastīdas. • Mitohondriji. • Viciņas un skropstiņas. • Citoskelets: mikrocaurulītes, mikrofilamenti, starpfilamenti.
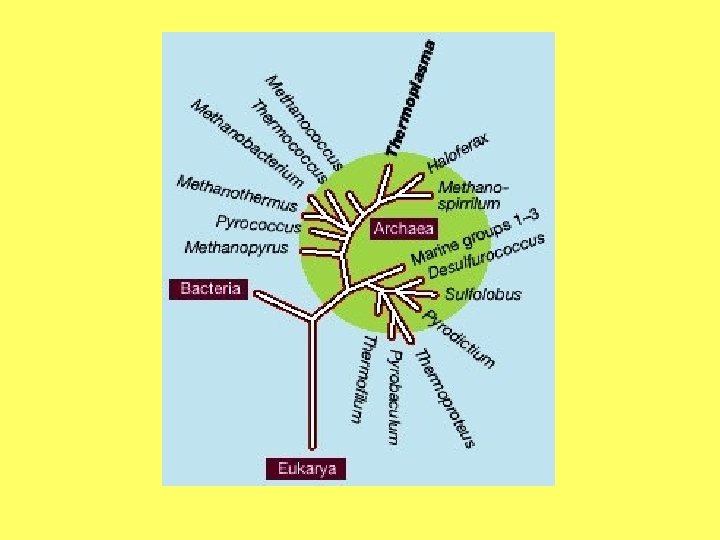
Endosimbiozes teorija • Prokariotiskā saimniekšūna ir bijusi līdzīga mūsdienu Thermoplasma, kura sastopama karstos un skābos ūdeņos (Margulis and Sagan 1987). • Mitohondriju un peroksisomu priekšteči ir mūsdienu Daptobacter un Bdellovibrio līdzīgi prokarioti, kuri efektīvi izmanto skābekli (Margulis and Sagans 1987). Peroksisomu čaperoni atgādina mitohondriālos un tiek kodēti kodolā. • Plastīdu priekšteči ir mūsdienu zilaļģēm Cyanobacteria līdzīgi organismi. • Eikariotiem tipisko viciņu priekšteči varētu būt mūsdienu Spirochetes līdzīgi organismi.
Eikarioti, kuriem nav mitohondriju un hlorplastu http: //www. ferris. edu/htmls/academics/course. offerings/clinlabs/courses/clls 225/PARSITOL/gltroimg. htm
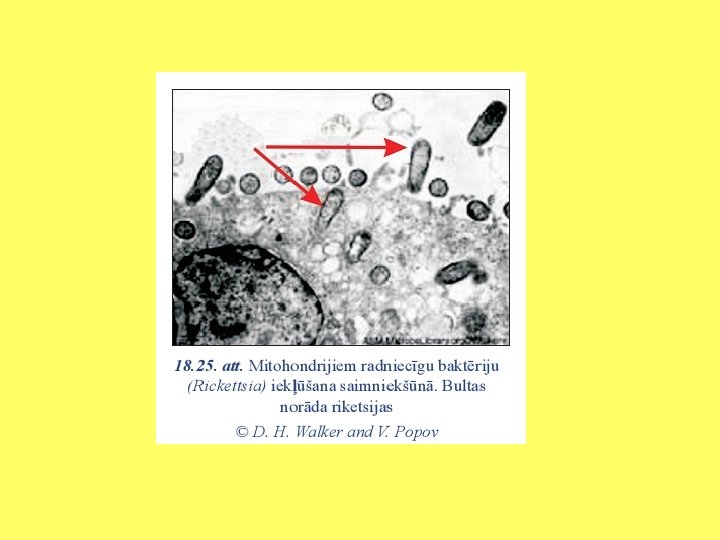
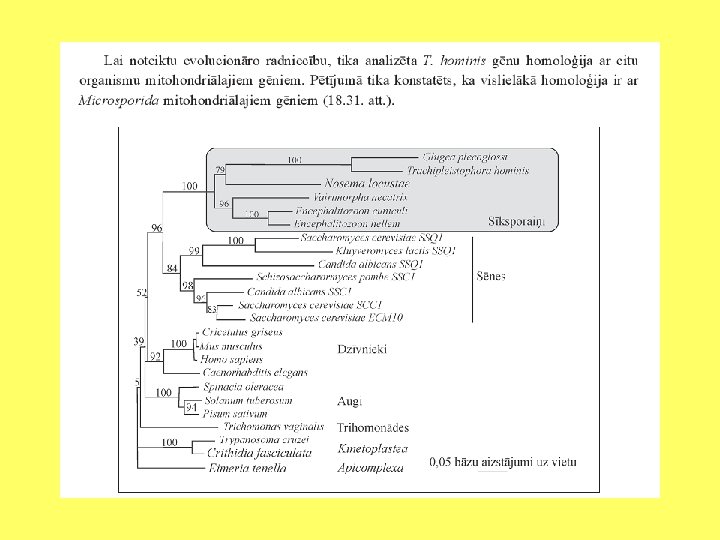
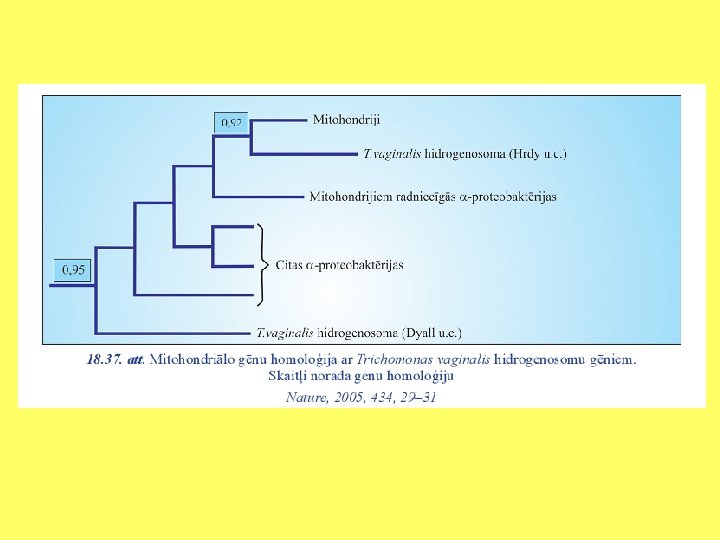
Hidrogenosomu un mitohondriju radniecība
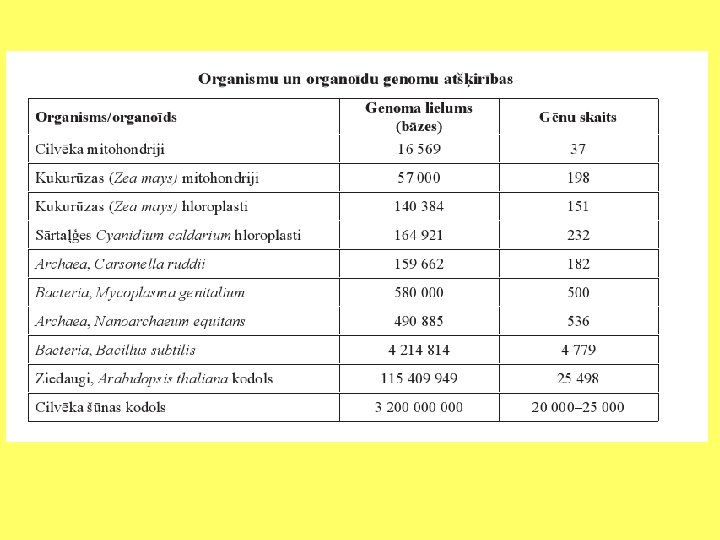
Analizēti gēni, kuri kodē elektronu pārnesi mitohondriju iekšējā membrānā un hidrogenosomās. Tika konstatēts, ka daudzi polipeptīdi, kurus kodē mitohondriālais genoms, ir atrodami hidrogenosomās.
Gēnu pārnese no organellām uz kodolu • Mitohondriji un hloroplasti kodē līdz 70 olbaltumvielām; • Peroksisomām nav DNS, bet vairojas daloties; • Centriolas vairojas daloties.
Gēnu transports no plastīdām un mitohondrijiem uz kodolu DNS uz kodolu var pārnest tieši vai ar RNS starpniecību. Raugu šūnās mitohondriju gēnu pārnese ir 1 uz 105 paaudzēm (Thorsness and Fox 1990 Blanchard and Schmidt 1995).
DNS pārnese baktēriju šūnās • konjugācija • transdukcija
DNS pārnese starp hloroplastiem Hloroplastu izaugumi (stromules). Tās savieno hloroplastus vienotā tīklā, transportē proteīnus, var transportēt pat DNS.
Gēnu transports no plastīdām un mitohondrijiem uz kodolu • Dinofītaļģēs (Heterocapsa triquetra) ir DNS minigredzeni 2 -3 kbp lieli. Katram ir konservatīvs nekodējošs reģions (Zhang et al. 1999). • Tos var viegli iesaiņot transportam un ar to izskaidro mazo hloroplastu genomu dinofītaļģēs (Mc. Fadden, 1999).
Gēnu transports no plastīdām un mitohondrijiem uz kodolu • Gēnu pārnešana var notikt arī bez transporta caur citosolu, jo šūnās iespējami kodola ārējās membrānas kontakti ar hloroplastu un mitohondriju ārējām membrānām.
Organellu ārējo membrānu kontakti
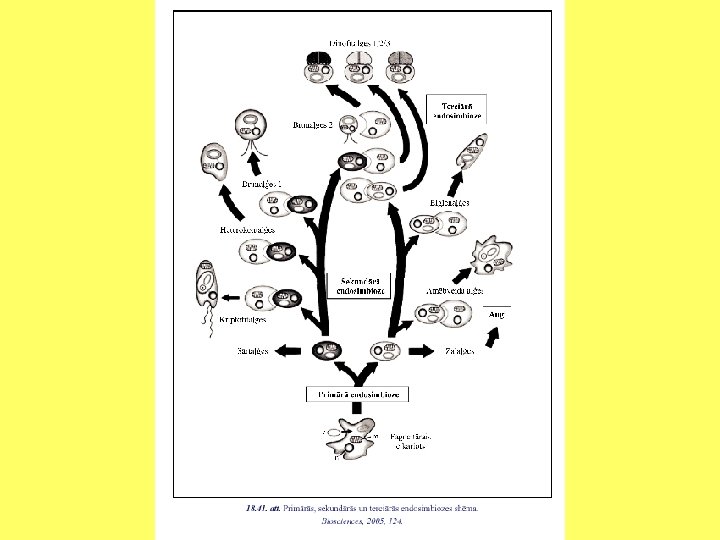
Sekundārā endosimbioze Daudzveidīgās plastīdas liek domāt paralēlu evolūciju vai savstarpēji atšķirīgu izcelsmi. Piecas galvenās grupas: • Rhodophyta - hlorofils a, 2 membrānas. • Heterokonts - hlorofils a un c, 4 membrānas. • Dinoflagellates - hlorofils a un c, 3 membrānas. • Euglenoids - hlorofils a un b, 3 membrānas. • Chlorophyta - hlorofils a un b, 2 membrānas.
Sekundārā endosimbioze Kriptomonādes - eikariotiskas, vicainas šūnas, kuras ar endocitozes palīdzību iekļāvušas sārtaļģes.
Sekundārā endosimbioze Nukleomorfs satur 3 hromosomas un kodē nedaudzu olbaltumvielu sintēzi.
Sekundārā endosimbioze