nu augana diferencicija un novecoana Starpnu tilti starp












 § Permanentie audi (skrimšļaudi)")

")
 Tābakas šūnu nāve. B – PCD")












- Slides: 29
Šūnu augšana diferenciācija un novecošana
Starpšūnu tilti starp ovocītiem un pavadītājšūnām http: //www. jcb. org/cgi/content/full/154/3/480 -a
Embrionālās attīstības sākums
Augļu mušiņu olās gēnam bcd - “bicoid” - veidojas m. RNS gradients. Ola saņem transkriptus no pavadītājšūnām. Transporta virziens un signāls RNS molekulas 3’ daļā nosaka augstu šo m. RNS molekulu koncentrāciju anteriorajā daļā un zemu – pretējā pusē.
Pēc apaugļošanās sākas translācija. Izveidotā olbaltumviela ir atbildīga par kāpura galvas veidošanos. Savukārt olas pretējā pusē uzkrājas “nanos” olbaltumvielas, kuras ir atbildīgas par astes daļas veidošanos.
Drozofilas embrionālā attīstība 4, 5 stundas, stadijas 1 -9. § m. RNS FISH analīze, 25% Drozofilas genoma 3370 gēni § § § 3214 gēni ekspresēti, 2198 nāk no mātšūnas, 504 ekspresēti zigotā, 388 ekspresēti zigotā un mātšūnās. Mātšūna dod 65% no zigotas m. RNS!
71% no gēniem ir noteikta lokalizācija.
Virspuse, 3 stadija. cno novietots šūnu perifērijā, anillin novietots gredzenveidā pie kodoliem. m. RNS-zaļa, kodols – sarkans.
http: //bio 1151. nicerweb. com/Locked/media/ch 18/bicoid 2. html
Makrofāgs Eritrocīts Cilmšūna kaula smadzenēs Segmentkodolainais granulocīts Megakariocīts ar trombocītiem
Šūnu specializācija daudzšūnu organismos § Apmēram 50 mitožu rezultātā izveidojas ap 230 šūnu veidiem; § Šūnas satur vienus un tos pašus gēnus, bet katrai šūnu grupai transkribē citus gēnus. http: www. ncbi. nim. nih. gov/About/primer/genetics_cell. html
Organoīdu kvantitatīvas atšķirības dažādos diferenciētu šūnu tipos Gludo muskuļu šūna. Leikocīts ar heparīna u. c. olbv. ieslēgumiem Atšķiras šūnu lielums, forma, kodolu lielums un forma, mitohondriju lielums un skaits, citoskeleta uzbūve, endoplazmatiskā tīkla lielums, ieslēgumi, u. c. morfoloģiskas pazīmes.
Audu reģenerācija § Dinamiskie audi (limforetikulārie audi, plakanais epitēlijs) § Permanentie audi (skrimšļaudi)
Epitēlijs kā diferenciācijas modelis Termināla diferenciācija Šūnu diferenciācija sākas ar postmitotisku keratinocītu, bet beidzas ar nedzīvu šūnu. Diferenciācijā ir sekojoši etapi: • Aktīva keratīnu un citu struktūras proteīnu sintēze. • Goldži aparāta aktivēšanās un citu organellu zudums. • Šūnas saplacināšanās un samazināšanās. • Metabolisma specializācija. • Šūnas membrānas caurlaidības palielināšanās. • Dehidratācija.
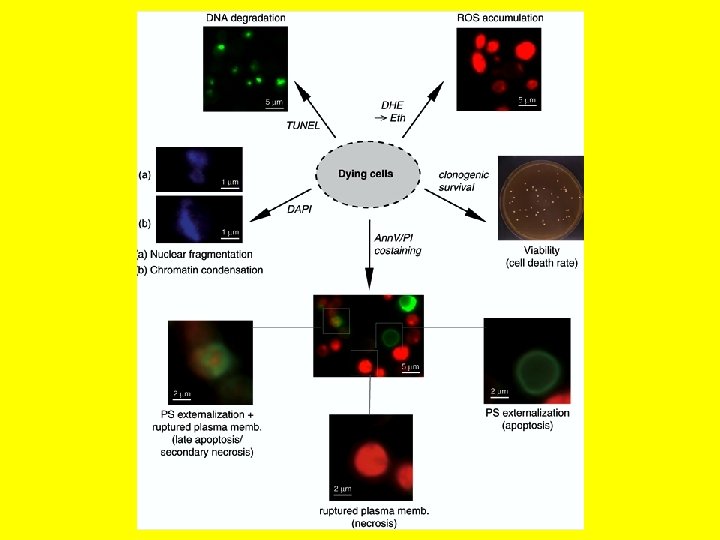
Šūnu nāve Visām šūnām var iestāties nāve. Tas notiek ievainojuma rezultātā - bojāeja (nekroze) vai pašnāvības rezultātā (programmēta šūnas nāve). § AUTOFĀGIJA/PROTEOSOMU-UBIKVITĪNA SISTĒMA/ § NEKROZE/PROGRAMMĒTA NĀVE
Journal of Cell Science 110, 1333 -1344 (1997) Tābakas šūnu nāve. B – PCD ierosinājis TMV, D – nekrozi izraisījusi sasaldēšana un atkausēšana.
Šūnu bojājumu cēloņi § Mehāniski bojājumi; § Fizikāli: temperatūra, UV un jonizējošā radiācija, mikroviļņi; § Ķīmiski: skābekļa savienojumi, brīvie radikāļi, genotoksiski un proteotoksiski savienojumi; § Barības vielu, skābekļa vai gaismas enerģijas trūkums; § Patogēni: vīrusi, baktērijas, vienšūņi un sēnes; § Programmēta šūnas nāve. § NEKROZE SAISTĪTA AR MEMBRĀNU BOJĀJUMIEM UN Ca 2+ IEKĻŪSANU CITOSOLĀ.
Ca 2+ koncentrācijas pieaugums izraisa nekrozi • Aktivē Ca 2+ atkarīgās fosfolipāzes (noārda membrānu lipīdus); • aktivē Ca 2+ atkarīgās proteāzes (noārda citoplazmas olbaltumvielas); • aktivē Ca 2+ atkarīgās endonukleāzes (noārda DNS un RNS); • noārda citoskeletu; • dzīvnieku šūnas fragmentējas; • augos saglabājas šūnu sieniņas.
Nekroze • • • Paplašinās kodola apvalka starpmembrānu telpa; kodola centrā kondensējas hromatīns; citoplazma kļūst tumšāka - olbaltumvielas noārdas (izšķīst ).
Programmētas šūnu nāves cēloņi § Katru DNS replikācijas reizi DNS saīsinās telomēras rajonā. DNS bojājumi aktivē programētu šūnu nāvi realizējošās olbaltumvielas. § Enzīms – telomerāze ir aktīvs tikai dažos šūnu tipos. Tajos DNS garums telomērās tiek atjaunots un šis programmētās šūnu nāves veids nevar notikt. § DNS bojājuma gadījumā aktivējas olbaltumviela p 53 un saistās ar bojāto DNS. § Aktivējās daudzi gēni un aktivē programmētu šūnu nāves kaskādi realizējošās olbaltumvielas. http: //en. wikipedia. org/wiki/Telomere
Programmēta šūnu nāve § § Šūnas izstiepjas; to mitohondriji sadalās, izdalot citohromu; ārpusē veido burbuļveida izaugumus; kodola DNS un olbaltumvielas fragmentējas, un veidojas mikrokodoli; § membrānu iekšienē novietotais fosfolipīds - fosfotidilserīns tiek pavērsts pret membrānas ārpusi; § pievienojas fagocitāro šūnu, piemēram, makrofāgu, membrānu receptoriem, un fagaocitozes ceļā šūnu fragmenti tiek sadalīti fagocītu lizosomās; § fagocitāro šūnu sekretētie citokīni kavē iekaisumu audos.
Apoptoze dzīvnieku šūnā noslēdzas ar šūnas fragmentāciju.
http: //www. apocyte. co. uk/apocyte/science/
Trofiskā faktora trūkuma gadījumā, pro apoptotiskā olbaltumviela Bad pievienojas pie anti apoptotiskām olbaltumvielām Bcl-2 un Bcl-xl, kuras atrodas mitohondriju membrānā. Tas traucē šīm olbaltumvielā saistīties ar membrānas olbaltumvielu Bax. Tā rezultātā Bax membrānā veido kanālus, kas pastiprina jonu plūsmu. Tas noved pie citohroma c atdalīšanās mitohondriju starpmembrānu telpā. Citohroms c savienojas ar adaptorolbaltumvielu Apaf-1. Tas izraisa kaspāžu aktivēšanas kaskādi, kas noved pie šūnas nāves. B. Pettmana un C. E. Hendersona (1998), Neuron 20, 633.
Trofiskā faktora klātbūtne vairākās šūnās aktivē proteīnkināzes Pl-3 aktivitāti. Tas aktivē proteīnkināzi Akt, kura fosforilē Bad. Fosforilētais Bad veido kompleksu ar olbaltumvielu 14 -33. Tas ļauj anti apoptotiskajām olbaltumvielām Bcl-2 un Bcl-xl inhibēt Bax aktivitāti. Tas novērš citohroma c izdalīšanos citosolā un neļauj iedarbināt kaspāžu aktivēšanas kaskādi. B. Pettmana un C. E. Hendersona (1998), Neuron 20, 633.
METODES DNS fragmentācija. Zirņu dīgsti 10 min turēti 55°C , lai inducētu PCD. DNS ir izolēta un atdalīta ar agarozes gela elektroforēzi. DNS pakāpeniska fragmentācija (laddering). Parādās kā 200 bp lielu fragmentu multimēri. Fragmenti rods tāpēc, ka starp nukleosomām tiek pārrauta DNS molekula. Kreisajā pusē molekulmasas marķieri.
Šūnu nāve rīsu suspensijas šūnās, kas apstrādātas ar PA. Šūnu nāve tika novērtēta pēc Evansa krāsas piesaistīšanās daudzuma. A – Šūnu nāve inducēta ar PA no 0, 1 – 5 mg ml-1. B – 1 mg ml-1 lielas PA koncentrācijas izraisītās šūnu nāves laika līkne. C – apoptozes noteikšana ar FACS. Rīsu protoplasti inkubēti 24 st PA 1 mg ml-1 lielā koncentrācijā un krāsoti ar propīdija jodīdu. Fluoresence rāda DNS saturu šūnā. Šūnas ar DNS daudzumu mazāku par haploīdu tika uzskatītas par apoptoiskām. Cell Research (2004); 14(1): 27 -33
TUNEL metode apoptozes noteikšanai TUNEL metode. Protoplasti tika pētīti pēc 48 stundu apstrādes ar normālu gaisu (a, b) vai gaisu ar pazeminātu skābekļa koncentrāciju (c, d). Šūnas tika krāsotas ar TUNEL reaģentu un HO 342. b un d attēli parāda visus kodolus, c – TUNEL pozitīvus kodolus. TUNEL reaction (Td. T-mediated d. UTP nick-end labelling) Xu et al. / J Zhejiang Univ SCI 2004 5(2): 137 -143