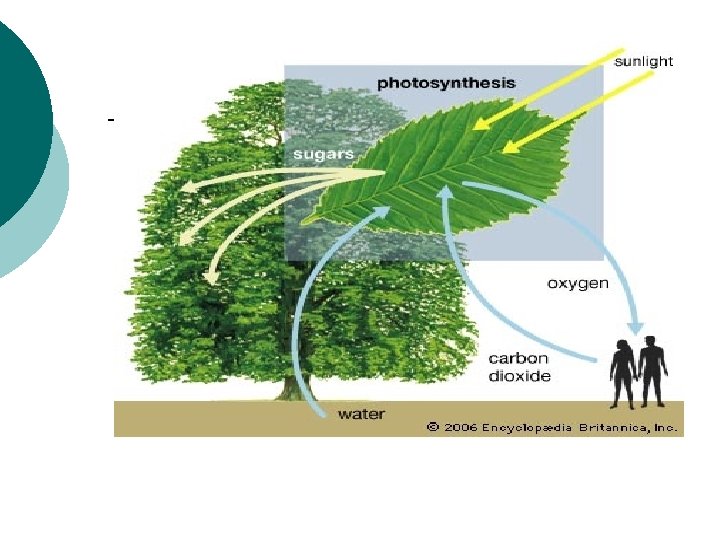
Znaaj svetlosne energije u procesu fotosinteze Energija sunca



















 njegova energija")

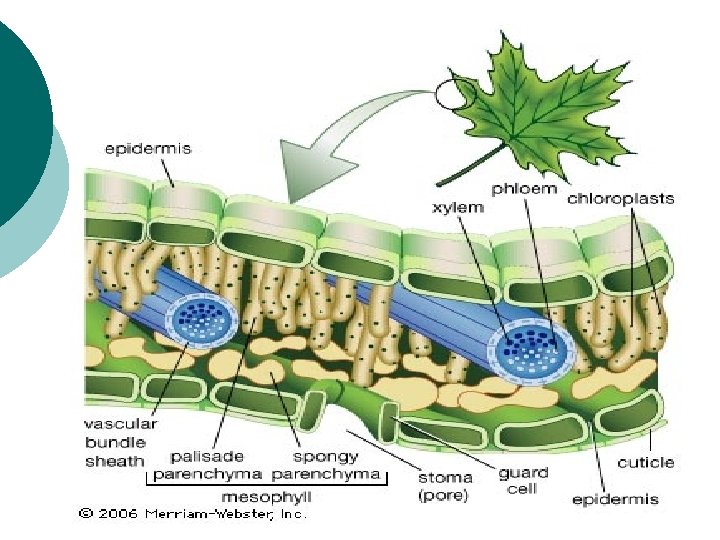
 ¡ ¡ U hloroplastima se na tilakoidima nalaze pigmenti i")

 ¡ ¡ Proizvodi svetle faze, ATP (sadrži energiju) i NADPH")


 ugradnjom")



- Slides: 41
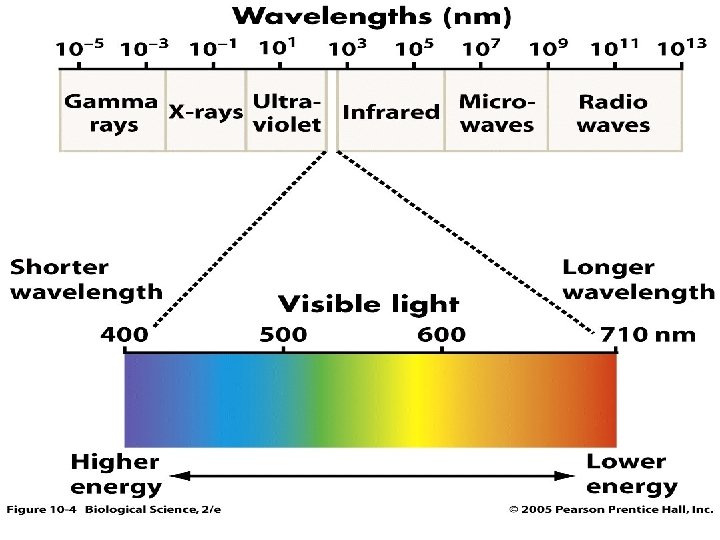
Značaj svetlosne energije u procesu fotosinteze -Energija sunca dospeva na zemlju u vidu elektromagnetnog zračenja. Sunčeva svetlost se sastoji od svetlosnih talasa različitih dužina koji nemaju isti značaj za fotosintezu. OSOBINE SUNČEVE SVETLOSTI, APSORCIJA I DEJSTVO SVETLOSNIH TALASA RAZLIČITE DUŽINE Biljke mogu da koriste direktnu i difuznu svetlost. Difuzno zračenje pri srednjim geografskim širinama je značajno (30 -40% od globalnog zračenja). Difuzno zračenje bolje prodire u donje delove useva nego direktno zračenje. Za životne procese biljaka najveći značaj ima integralna, kratkotalasna radijacija koja se uslovno može deliti na: 1. Ultaljubičasto zračenje (od 300 -390 nm). . . . 9% 2. Vidljivi deo spektra ( od 390 -760 nm). . . . 40% 3. Bliska- infracrvena radijacija (preko 760 nm). . . . . 51% Ako se imaju u vidu fototrofne bakterije tada za biljke značajni elektromagnetni spektar obuhvata oblast od 300 do 1100 nm.
FOTOSINTETIČKI AKTIVNA RADIJACIJA-ona se u viših biljaka kreće od 390 -710 nm, purpurne bakterije 300 -950 nm. -Zračenje iznad 1100 nm ne izaziva pobuđivanje elektrona (kvanti imaju previše malu energiju). Fotohemijske reakcije započinju samo onda kada energija kvanta date svetlosti nije manja od energije koja je potrebna da počne fotohemijska reakcija Ultraljubičasto zračenje kraćih talasnih dužina od 290 nm deluju destruktivno na žive organizme(apsorbuje ih sloj ozona koji se nalazi na visini između 15 -30 km. ) Ukoliko je talasna dužina svetlosti manja utoliko je njena energija veća i obrnuto. Fotosintetički aktivna radijacija se: 1. Reflektuje. . . . 5 -12% 2. Prolazi kroz tkivo biljaka. . . . . 5 -8 % 3. Apsorbuje i koristi za fiziološke procese. . . 70 -90 % Blisku infracrvenu svetlost apsorbuje voda koja se nalazi u biljkama. Biljke u toku vegetacije prosečno iskorišćavaju sunčevu energiju sa svega 1 -2 % (mala asimilaciona površina biljaka na početku i kraju vegetacije. )
Uticaj intenziteta svetlosti na fotosintezu Heliofite-vrste koje zahtevaju punu osvetljenost. Neto fotosinteza se dugo povećava sa povećanjem intenziteta osvetljenosti. Skiofite-ne podnose jako osvetljenje i izdržavaju zasenjivanje (spadaju i korovske biljke). Kompenzaciona tačka za svetlost postiže već pri relativno maloj osvetljenosti. Kompenzaciona tačka za svetlost -predstavlja intenzitet svetlosti pri kojem uz nepromenjivu koncentraciju CO 2 dolazi do ravnoteže između količine fotosintezom usvojenog i disanjem odatog CO 2. -ukazuje na potrebu određene biljke za svetlošću -važna je za useve sa gustim sklopom biljaka. Mladi listovi se mogu u određenim granicama prilagođavati visokom ili niskom intenzitetu osvetljenosti. Prilagođavanje fotosintetičkog aparata na uslove različite osvetljenosti naročito je značajno kod biljaka koje se rasađuju.
Uticaj spektralnog sastava svetlosti na fotosintezu -od posebnog je značaja pri gajenju biljaka u veštačkim uslovima - Pri niskom položaju Sunca put svetlosnih zraka u atmosferi je duži, usled čega se zraci kraćih talasnih dužina više rasejavaju, tako da na Zemljinu površinu dospeva više crvenih zrakova i obrnuto, ukoliko je ugao veći sve je više zrakova kratkih talasnih dužina. - U voćnjacima pravac redova treba da bude severjug, tada će stabla biti najbolje izložena svetlosti - Spektralni sastav utiče na: veličinu hloroplasta, brzinu formiranja njihove strukture, broj i strukturu tilakoida. . .
Uticaj koncentracije CO 2 na fotosintezu Koncentracija CO 2 U donjim slojevima atmosfere po zapremini u proseku iznosi 0, 03%(od o, o 25 –o, o 35). Koncentracija CO 2 u gornjim slojevima atmosfere je manja. Smanjenje koncentracije CO 2 za svega 15 -20% od normalne vrednosti može značajno da smanji intenzitet fotosinteze, a da pri koncentracijama nanjim od 0, 010 do 0, 008% dolazi do depresije fotosinteze. Intenzitet fotosinteze se povećava sa povećanjem koncentracije CO 2 u atmosferi. Koncentracija CO 2 iznad koje se intenzitet fotosinteze više ne povećava naziva se saturaciona ili zasićujuća koncentracija pošto zasićuje fotosintetički aparat(0, 18 – 0, 35% CO 2) Pri gajenju biljaka u zatvorenom prostoru moguće je povećanjem koncentracije CO 2 u atmosferi povećati prinos 20 i više puta. Povećanje koncentracije CO 2 u atmosferi ponekad se naziva i “ đubrenje sa CO 2 “. U praksi potrebno je upotrebom organskih đubriva i obradom povećati mikrobiološku aktivnost i time i “ disanje zemljišta “.
Uticaj koncentracije kiseonika Neto fotosinteza je u anaerobnim uslovima određeno vreme 1, 5 -2 x veća nego u atmosferi sa normalnim sadržajem kiseonika (Warburgov-efekat) Warburgov-efekat predstavlja stimulaciju fotorespiracije prisustvom kiseonika na račun fotosinteze. O 2 je supstrat glikolatfosfat-sintetaze i kompetitivno inhibira fiksaciju CO 2 preko ribulozebisfosfatkarboksilaze. Aerobni uslovi podstiču fotorespiraciju i time razgradnju organske materije. Svaki 4 atom C-ka koji se uključuje u fotorespiraciju izlučuje se u vidu CO 2. Smanjenjem koncentracije O 2 neto fotosinteza može se povećati za 50 -100%, zahvaljujući inhibiciji reakcija fotorespiracije u kojima se troši kiseonik. Nepovoljno dejstvo kiseonika na neto fotosintezu može se ublažiti smanjenjem t-re i povećanjem koncetracije CO 2.
Uticaj temperature na fotosintezu Razlika između temperature vazduha koji okružuje listove i samih listova može da bude i do ± 10° C. Na svetlu fazu fotosinteze ne utiče temperatura u okviru bioloških granica pošto je ona fotohemijske prirode. Tamna faza – koja se sastoji od niza biohemijskih reakcija jako je zavisna od temperature. U uslovima dovoljno visoke koncentracije CO 2 i pri zasićenju listova svetlošću, bruto fotosinteza povećanjem temperature raste praktično sve dok ne dođe do inaktivacije enzimskog sistema fotosintetičkig aparata. Temperatura može direktno(preko nivoa respiracije) i indirektno(zatvaranje stoma) da utiče na fotosintezu.
Uticaj mineralne ishrane na fotosintezu Direktan uticaj neophodnih elemenata može se prepoznati brzim dejstvom, već posle nekoliko minuta ili sati nakon njihovog dodavanja biljkama kojima nedostaju (Mn, fosfatni joni, joni kalijuma. ) Indirektno dejstvo se pripisuje N, Fe, Mg, S i dr. -Mn i Cl se pripisuje uloga pri fotooksidaciji vode. Za P se smatra da utiče na tamne reakcije fotosinteze. Njegov nedostatak inhibira redukcione procese fotosinteze, iako je on neophodan i u svetloj fazi pri fotofosforilaciji. - K utiče na aktivnost brojnih enzima , na strukturu hloroplasta, učestvuje u regulaciji pokreta ćelija zatvaračica stominog aparata a time i na usvajanje CO 2. N – ulazi u sastav mnogih organskih jedinjenja Količinu i aktivnost ključnih enzima fotosinteze Povoljno utiče na obrazovanje lisne površine, dužinu života listova, sadržaj hlorofila Mg – sastav hlorofila Neki metali ulaze u sastav komponenata elektrotransportnog lanca Fe(feredoksin i citohrome), Cu (plastocijanin)
BIOENERGETSKI I GLOBALNI ZNAČAJ FOTOSINTEZE - - - Procesom fotosinteze biljke svetlosnu energiju preobraćaju u energiju makroenergetskih jedinjenja koju zatim koriste za sintezu organskih materija. U CO 2, supstratu fotosinteze ugljenik je maksimalno oksidisan Fotosinteza ( najuopštenije) je redoks proces u kome sa donora elektrona, elektron prelazi na akceptor elektrona, pri čemu energiju za savladavanje razlike u redokspotencijalu daje svetlost - Više biljke kao donor elektrona, odnosno vodonika za redukciju CO 2 koriste vodu hv H 2 D + A → H 2 A + D hv 6 CO 2 + 12 H 2 O → 6(CH 20 ) + 6 H 2 O +6 O 2 hlorofil ΔG=+2850 KJ molˉ¹ (nivo transformacije svetlosne energije)
Efekti fotosintetičke aktivnosti -količina ugljenika koja se godišnje vezuje u organska jedinjenja na Zemlji iznosi 44 milijarde tona. Vezuje se energija i produkuje organska materija Pri usvajanju svake 1 t C oslobađa se 2, 7 t O 2. Fotosinteza je omogućila pojavu aerobnog disanja. Kiseonik atmosfere interaguje sa ultraljubičastim zracima koji dolaze od Sunca. Fotosinteza omogućava, a istovremeno je i zavisna od permanentnog kruženja elemenata u prirodi. Pošto su za sintezu organske materije osim C, O i H potrebni i drugi elementi ( N, P, S i dr. ), zahvaljujući fotosintezi dolazi do njihovog kruženja u prirodi. INTENZITET FOTOSINTEZE – količina usvojenog CO 2 ili izdvojenog O 2 na jedinicu lisne površine ili mase za jedinicu vremena. Izražava se u mg CO 2 ·dm ˉ² ·h ˉ¹, odnosno mg CO 2 ·g ˉ¹ sveže ili suve materije ·h ˉ¹. PRODUKTIVNOST FOTOSINTEZE – količina stvorene organske materije po jedinici lisne mase u jedinici vremena. Izražava u g ·dm ˉ² · dan ˉ¹.
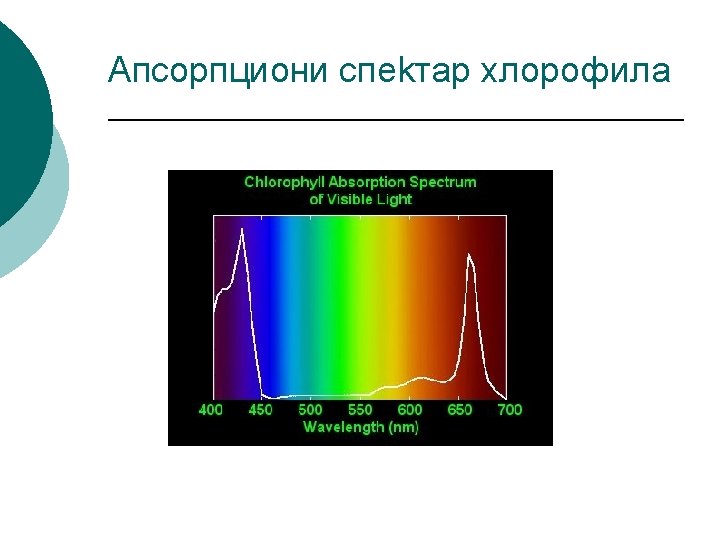
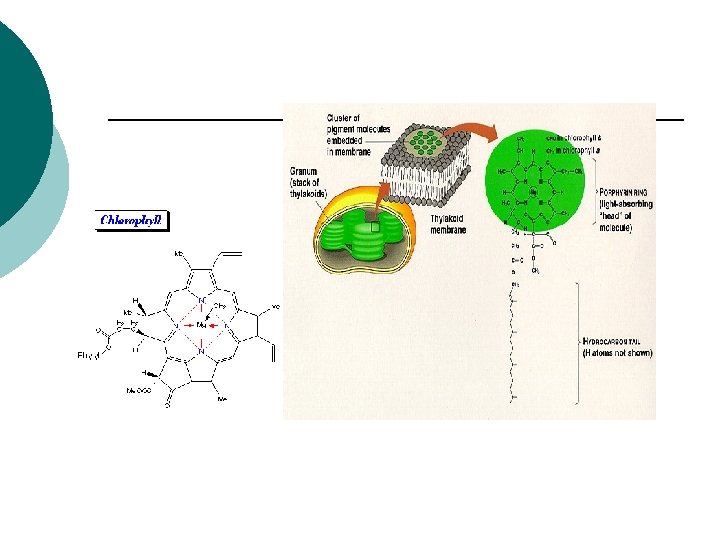
Fotosintetski pigmenti -U prirodi se nalaze 2 grupe jedinjenja koja apsorbuju svetlost, to su tetrapiroli (prstenasti grade hlorofili) i izopreni ( grade karotenoide). -Linearni tetrapirol nalazi se u fitohromima. -Deo vidljivog dela spektra sunčeve svetlosti biljke mogu pomoću fotosintetičkih pigmenata, fotoreceptora da apsorbuju i da koriste u fotosintezi. -Apsorcija svetlosti vidljivog dela spektra i s tim u vezi boja fotosintetičkih pigmenata zasniva se na prisustvu sistema konjugovanih dvostrukih veza u njihovim molekulima : --CH=CH-CH=CH ili -CH=CH-CH=N-CH=CH ili , sličnih nezasićenih grupa -CH =N-, -N =O, =C =S -NAJZNAČAJNIJI PIGMENTI SU : HLOROFIL I KAROTENOIDI
HLOROFIL – HEMIJSKE I FIZIČKE OSOBINE ¡ ¡ ¡ ¡ Hlorofil a C 55 H 72 O 5 N 4 Mg (oblici P 700 i P 680) Hlorofil b C 55 H 70 O 6 N 4 Mg Hlorofil a i Hlorofil b su estri dikarbonske kiseline hlorofilina, gde je jedan vodonik na jednoj –COOH grupi zamenjen ostatkom alkohola fitola, a u drugoj ostatkom metil-alkohola. Osnovna gradivna jedinica hlorofila su četri pirolova petočlana prstena koji su međusobno povezani metinskim grupama -CH= čineći na taj način porfirinski prsten U središtu porfirinskog prstena nalazi se Mg koji je vezan za atome azota pirolovog prstena Hlorofil b se razlikuje od hlorofila a samo po tome što se na trećem ugljenikovom atomu drugog pirolovog prstena umesto metil (CH 3) nalazni aldehidna grupa (COH) Odnos Hlorofil a : Hlorofil b = 3 : 1 Hlorofil b veoma efikasno prenosi sopstvenu energiju iz pobuđenog stanja na hlorofil a.
Hlorofil-protein kompleksi 1. 2. 3. 4. Fotosintetski pigmenti svoju specifičnu ulogu mogu da obave samo ako su vezani za određene strukturne elemente ćelije. Ako se veza prekine, prestaje biohemijska aktivnost pigmenata. Dok je fitolni rep u vodi nerastvorljiv(hidrofoban), dotle je porfirinsko jezgro, a posebno izocikličan pentanon prsten u vodi rastvorljiv (hidrofilan). Molekuli hlorofila mogu da se vežu sa drugim jedinjenjima koja imaju takođe i hidrofilne i hidrofobne osobine. Postojanje hlorofil-protein kompleksa dokazuje različiti spektar apsorcije izolovanog i za ćeliju u tilakoidima hloroplasta vezanog pigmenta Ako se vodenom ekstraktu hlorofila dodaje aceton, protein se taloži i pigment prelazi u rastvor. Ova pojava ukazuje da veza između pigmenta i proteina nije naročito jaka.
Činioci koji utiču na sadržaj hlorofila 1. 2. 3. 4. 5. 6. Svetlost utiče na sadržaj hlorofila svojim kvalitetom i intenzitetom. Intenzitet ozelenjavanja najveći pri crvenoj, zatim plavoj a najmanji pri zelenoj svetlosti. Zasenjeni listovi, i biljke osvetljene samo difuznom svetlošću, obično imaju veći sadržaj hlorofila od listova koji su izloženi direktnoj svetlosti. Pri većem intenzitetu svetlosti ravnoteža između sinteze i razgradnje uspostavlja se pri nižim koncentracijama hlorofila. Kardinalne temperaturne tačke za sintezu hlorofila (MIN 1 -2°C, MAX 40°C) optimum-25 °C Elementi: Mg, N, Fe Koncentracija O 2, Vlažnost zemljišta i vazduha, zagađenje (pesticidi, teški metali, štetni gasovi-SO 2 i dr. ) Unutrašnji činioci (botanička pripadnost, starost, zdravstveno stanje)
Karotenoidi- hemijske osobine i rasprostranjenost 1. 2. 3. 4. 5. 6. 7. Karotenoidi su produkti izoprena CH 2=C-CH = CH 2 | CH 3 (izopren) Njihov uglenikov lanac izgrađen je od 8 izoprenskih jedinica. Karotenoidi mogu biti bez, sa jednim, ili dva šestočlana prstena jonona. Zbog velikog broja konjugovanih dvostrukih veza oni su obojeni, a zbog dugog lanca ugljovodonika rastvorljivi su u lipoidima. Karotenoidi se dele na: karotene i ksantofile Karoteni su ugljovodonici naranđžastocrvene boje, a ksantofili sadrže kiseonik u hidroksi-, keto-, ili metoksigrupi i žute su boje Od ukupne količine karotenoida u proseku 20 -40% otpada na karotene, od čega preko 70 % na β-karoten, a 60 -80% na ksantofile: lutein, violaksantin, zeaksantin.
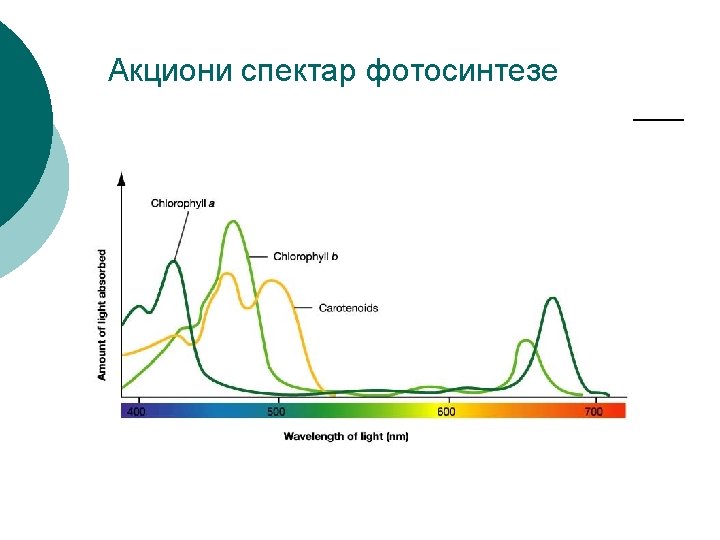
Fiziološka uloga karotenoida 1. 2. 3. 4. 5. 6. 7. Prenose energiju apsorbovanih kvanata na hlorofil a. Proširuju spektar apsorcije fotosintetičkog aparata Apsorbuju 10 -20% ukupne Sunčeve svetlosti Smatra se da fukoksantin, a možda i β-karoten, mogu da prenose apsorbovanu svetlosnu energiju bez gubitka na hlorofil a, dok je efikasnost drugih karotenoida znatno manja, posebno je niska u ksantofila. Štite fotolabilan fotosintetički sistem od oksidativne fotodestrukcije (učestvuju u stabilizaciji protein kompleksa). Pošto je β-karoten provitamin A, njegov sadržaj je značajan za biološku vrednost biljnih proizvoda Utiču na izgled i tržišnu vrednost biljnih proizvoda
Mehanizam fotosinteze Dve faze: svetla i tamna faza fotosinteze U svetloj fazi se stvaraju neophodni preduslovi za redukciju CO 2: redukcioni ekvivalenti i energija. Ona obuhvata sledeće etape: Apsorciju i migraciju svetlosne energije Razgradnju vode i obrazovanje O 2 Stvaranje redukcionih ekvivalenata – NADPH +H Fotofosforilacija- sinteza ATP Tamna faza obuhvata fiksaciju CO 2 i njegovu redukciju pomoću NADPH +H i ATP koji su stvoreni u svetlosnoj fazi. Za odvijanje ove faze nije potrebna svetlost, mada je ona potrebna za aktivaciju nekih enzima Calvinovog ciklusa.
Pobuđivanje molekula hlorofila -Kada jedan molekul hlorofila apsorbuje jedan kvant svetlosti (foton) njegova energija će se povećati. Energijom obogaćen molekul prelazi u ekscitirano stanje -kada se molekul vraća u osnovno stanje on odaje apsorbovanu energiju. Fotoreceptori nisu u stanju da prime novi kvant svetlosti sve dotle dok se ne vrate u osnovno stanje. - Pobuđeno stanje→jedan elektron prelazi na orbitalu veće energije - Za fotosintezu je naročito značajno pobuđeno singletno stanje, pošto se apsorbovana svetlosna energija u fotosintetičkom aparatu prenosi takvim stanjem preko pojedinih oblika hlorofila do reakcionog centra.
Vraćanje molekula u osnovno stanje -molekul za veoma kratko vreme gubi višak energije i prelazi u osnovno stanje. Molekule hlorofila a prvenstveno pobuđuje plava i crvena svetlost. Pri prelazu sa prvog pobuđenog singletnog stanja u osnovno stanje S 1→S 0 višak energije može da se oslobodi u različitim oblicima: -u vidu toplote -emisijom kvanta svetlosti-fluorescencija (3 -6% od ukupne a. e. ) -prenosom energije na susedni molekul bez zračenja, od molekula do molekula, do jednog reakcionog centra - fotohemijski rad-prenos elektrona nasuprot pada gradijenta redokspotencijala. -u membranama tilakoida u antena pigmentima dominira prenos energije, a u reakcionim centrima fotohemijski rad.
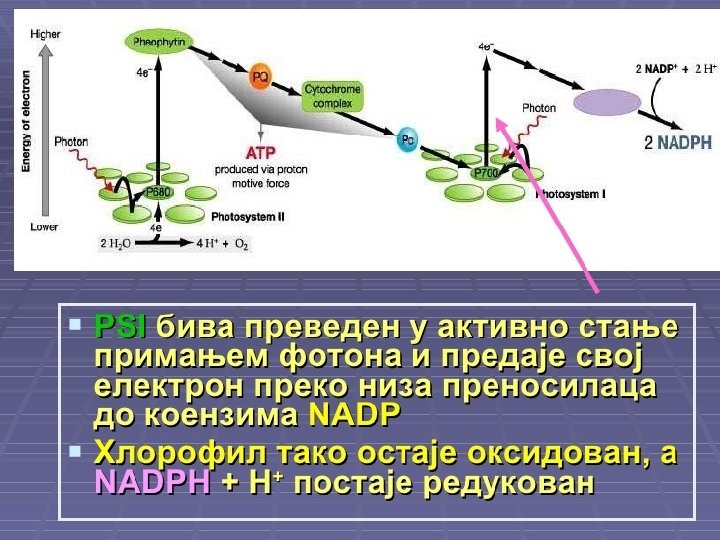
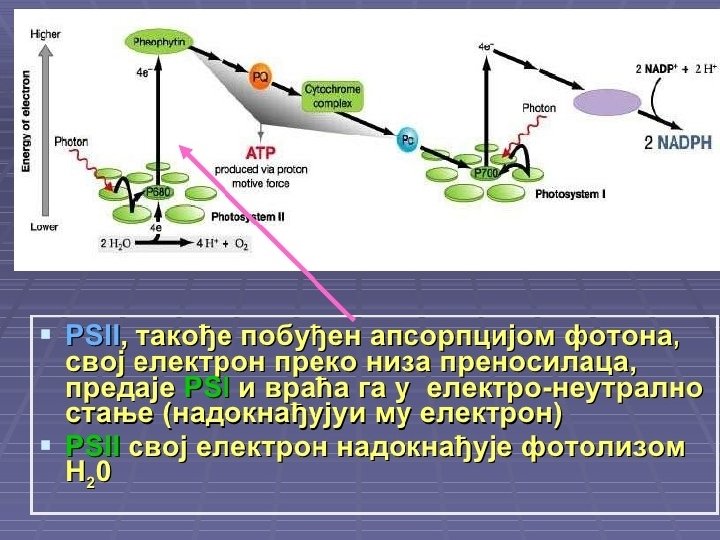
Svetla faza (fotohemijska faza) ¡ ¡ U hloroplastima se na tilakoidima nalaze pigmenti i enzimi koji zajedno nagrade dva fotosistema: FS 1 i FS 2. Kada molekul hlorofila apsorbuje svetlost, njegov elektron na spoljnoj orbitali postaje pobuđen, usled viška energije, pa napušta molekul hlorofila. Oslobođeni elektron prihvataju prenosioci (transportni lanac elektrona), koji su poređani tako da elektron uvek sa višeg prelazi na niži energetski nivo. Prelaskom sa višeg na niži energetski nivo elektron otpušta deo energije koju prima ADP-a i pretvara se u ATP predstavlja glavni izvor energije u ćeliji za sve njene funkcije. Svetlosna energija, pretvorena u električnu (energija elektrona) je krajnje transformisana u korisnu hemijsku energiju.
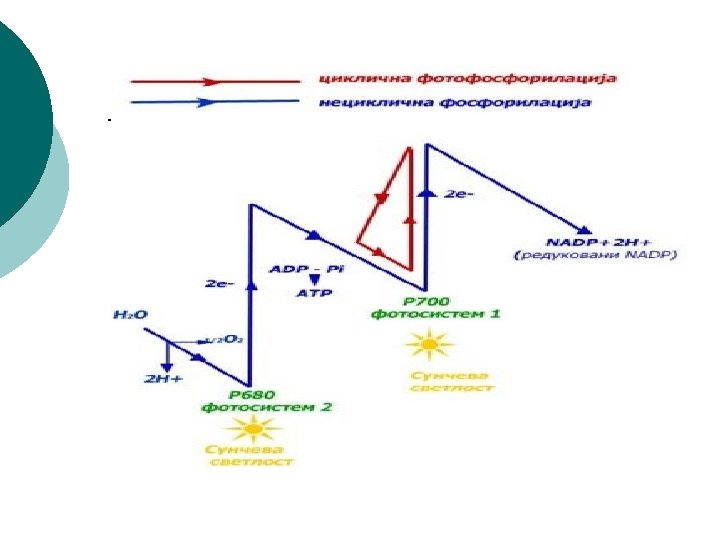
Sinteza ATP u svetloj fazi fotosintezi naziva se fotofosforilacija i može biti: ¡ ¡ ¡ ¡ ciklična, u kojoj elektron izbačen iz fotosistema 1 preko niza prenosilaca (transportni lanac elektrona) ponovo vraća u FS 1; u ovom procesu ne učestvuje NADP; neciklična U necikličnoj fosforilaciji krajnji primalac elektrona je koenzim NADP koji primanjem elektrona postaje redukovani NADPH 2. Fotosistem 1 svoj izgubljeni elektron nadoknađuje iz FS 2, a FS 2 nadoknađuje elektron iz vode. Voda se razlaže na kiseonik (odlazi u atmosferu) i vodonikove jone koje prihvata NADP i postaje NADPH 2. Prema tome, voda je primarni davalac, a NADP krajnji primalac elektrona u svetloj fazi. Krajnji proizvodi svetle faze su: kiseonik ATP i redukovani NADPH 2. ATP i NADPH 2 odlaze u tamnu fazu, a kiseonik se ispušta u atmosferu.
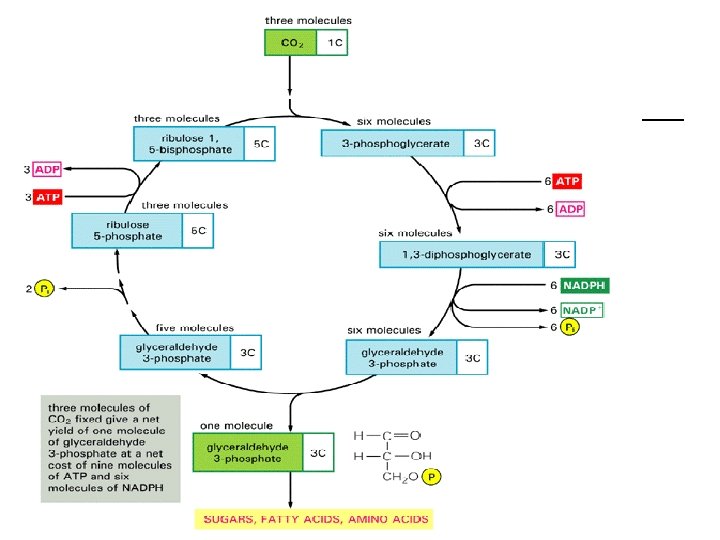
Tamna faza (termohemijska faza) ¡ ¡ Proizvodi svetle faze, ATP (sadrži energiju) i NADPH 2 (donosi vodonik poreklom iz vode), se koriste u tamnoj fazi da bi se neorganski ugljenik iz CO 2 ugradio u organska jedinjenja. To se naziva fiksacija ugljenika i izvodi se u Kalvinovom ciklusu u stromi hloroplasta. Početno jedinjenje ovog ciklusa je istovremeno i završno i naziva se ribulozodifosfat (Ru. DP). Ugljen-dioksid iz atmosfere ulazi u ćeliju (hloroplaste) i vezuje se za Ru. DP (ima 5 C atoma) pri čemu nastaje jedno nestabilno jedinjenje sa 6 C atoma koje se razlaže na dva molekula fosfoglicerinske kiseline (sa po tri C atoma). Da je to prvi stabilan proizvod tamne faze fotosinteze otkrio je Kalvin i za to otkriće 1961. g. dobio Nobelovu nagradu za hemiju. Preko niza proizvoda u toku ciklusa opet se stvara Ru. DP. Pri okretanju jednog Kalvinovog ciklusa u organska jedinjenja se ugradi jedan C atoma iz CO 2. Znači, da bi nastao jedan molekul fruktoze ciklus mora da se okrene 6 puta.
KALVINOV CIKLUS ¡ ¡ ¡ Ovaj ciklus je dobio ime po naučniku koji je sa saradnicima utvrdio raspored biohemijskih reakcija u tamnoj fazi i otkrio njene produkte. Za ova istraživanja Kalvin je dobio Nobelovu nagradu 1961. god. za hemiju. Proces se odvija tako što ugljen-dioksid koji je ušao kroz stome u ćelije mezofila vezuje se za organsko jedinjenje ribulozu 1, 5 - bisfosfat ( Ru. BP). Ta hemijsaka reakcija vezivanaja ugljendioksida naziva se karboksilacija , a enzim koji vezuje CO 2 je Ru. BP karboksilaza. Dobijeno jedinjenje ima šest C-atoma nestabilno je i brzo se razlaže na dva molekula 3 -fosfoglicerinske kiseline ( sa tri Catoma). Fosfoglicerinska kiselina je prvi stabilan produkt fotosinteze. Dalje reakcije se odigravaju uz učešće produkata svijetle faze fotosinteze. Uz učešće ATP, fosfoglicerinska kiselina se fosforiluje u 1, 3 fosfglicerinsku kiselinu, a zatim se pomoću NADPH redukuje ( od kiselinske grupe COOH postaje aldehidna. CHO) i dobija se gliceraldehid-3 -fosfat. On se pretvara u dihidroksiacetonfosfat. Spajanjem ove dvije trioze dobija se šećer sa 6 Catoma, fruktoza 1, 6 -bisfosfat.
KALVINOV CIKLUS ¡ ¡ Ovo su najvažnije reakcije u Kalvinovom ciklusu. U daljem toku ciklusa dijelovi ugljeničnog lanca od 6 C-atoma kombinuju se sa triozom na različite načine, dajući šećere sa 4, 5 i 7 ugljenika. Taj dio ciklusa se zove konverzija ( pretvaranje) šećera, da bi se ponovo regenerisalaribulozafosfat. Uz učešće još jednog molekula ATP, ovaj šećer se fosforiluje i postaje ribuloza 1, 5 -bisfosfat koja može primiti sledeći molekul CO 2. Pri svakom obrtaju ciklusa ugradi se po jedan Catom u organsko jedinjenje, a zatim se izvrši njegova redukcija i tako se ugljenik iz neorganskog oblika prevodi u organski. Da bi se dobio jedan molekul fruktoze potrebno je da se ciklus ponovi šest puta. Različiti šećeri nastali u Kalvinovom ciklusu ( eritroza, sedoheptuloza, pentoze) služe kao početni produkti za sinteze složenih organskih jedinjenja. Sekundarni produkti fotosinteze: skrob, saharoza, lipidi, organske kiseline, aminokiseline stvaraju se u hloroplastima ili citoplazmi u ćelijama mezofila lista odakle se transpotuju u druge dijelove biljke.
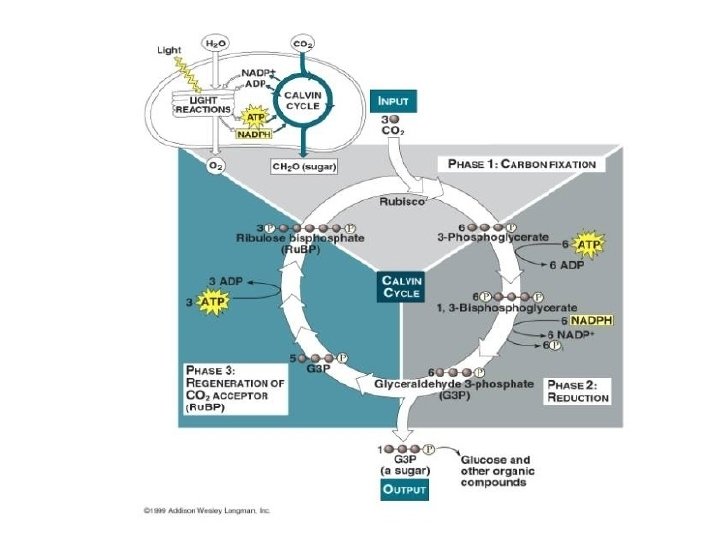
KALVINOV CIKLUS tri faze: • fiksacija CO 2 (akceptor ribuloza 1 -5 bifosfat) ugradnjom CO 2 stvara se heksoza koja se raspada na dvije molekule 3 - fosfoglicerata • redukcija 3 - fosfoglicerat se uz pomoć ATP-a i NADPH 2 reducira u 3 - fosfogliceraldehid je osnovno jedinjenje pri nastajanju šećera • regeneracija akceptora CO 2, ribuloze 1 -5 bifosfata ponovo se omogućava fiksacija C 02, a time i obnavljanje ciklusa
KALVINOV CIKLUS fiksacija CO 2 • početak Kalvinovog ciklusa vezan je uz fiksaciju ugljik - dioksida • akceptor je ribuloza 1 -5 bifosfat - reakciju katalizira enzim ribuloza 1 -5 bifosfat karboksilaza (Rubisco) • karboksilacijom nastaje heksoza koja se raspada na dvije molekule 3 - fosfoglicerata CO 2 + RUBISCO ENZIM 2
KALVINOV CIKLUS redukcija 3 - fosfoglicerata • 3 - fosfoglicerat se prvo uz pomoć ATP-a fosforilira do 1, 3 difosfoglicerata • reakciju katalizira enzim 3 - fosfoglicerat kinaza 3 – FOSFOGLICERAT KINAZA + ATP + ADP • dobiveni 1, 3 bifosfoglicerat se dehidrogenira u 3 - fosfogliceraldehid • reakciju katalizira enzim 3 - fosfoglicerat dehidrogenaza 3 – FOSFOGLICERAT DEHIDROGENAZA + NADPH 2 + NADP+
KALVINOV CIKLUS regeneracija akceptora CO 2, ribuloze 1 -5 bifosfata • molekule 3 - fosfoglicerata ulaze u brojne reakcije pri čemu nastaju šećeri • istovremeno jedan dio nastalih molekula 3 - fosfoglicerata se koristi za regeneraciju ribuloze 1 -5 bifosfata, spoja neophodnog za fiksaciju CO 2, čime se omogućava obnavljanje ciklusa