Zklady Genetiky Dag Lindgren Tom Funda ROSTLINN aaaa

 aaaaaaaa plastid ribozómy aaaaaaaa mitochondrie")





 22")






 Nicotina")
,")


 matrice DNA do m-RNA vzniká primární transkript m -RNA, následně podléhá")
 = skupina tří sousedních nukleotidů na mediátorové (informační) RNA, které svým")

 Translace = překlad proces překladu genetické informace z jazyka nukleotidů (kodonů) do pořadí")







 homologické chromozómy sesterské chromatidy po replikaci aaaaaaa segregace chromozómů")


 po rozdělení centromery putují k pólům pohyb je")




 se řadí v rovníkové rovině Anafáze I")
 Homeotypické dělení (meióza II) do značné míry podobné mitóze v anafázi II se")






 Ø je samosprašný Dominantní homozygot")









 podílí svým účinkem na realizaci")
- Slides: 57
Základy Genetiky Dag Lindgren Tomáš Funda
ROSTLINNÁ aaaa jádro BUŇKA chromatin (DNA) aaaaaaaa plastid ribozómy aaaaaaaa mitochondrie
Buňka Cell jádro chromozómy ribozómy aaaaaaa mitochondrie aaaaaaa jádroe eeeeee
Chromozóm Jádro Chromozómy Chromosome Chromatidy Telomera Centromera Buňka Jaderné Nukleotidové báze Telomera Histony Nukl DNA (dvojšroubovice)
Chromozómy Ø Vyskytují se v sadách Ø Nejčastější jsou diploidní organismy s 2 sadami, vyskytují se tedy v párech Ø Gamety (pohlavní buňky) mají pouze 1 sadu
Základní počty chromozómů Rod / Druh Betula pendula Populus tremula Salix Picea abies Pinus silvestris Sequoia sempervirens Tilia Taxus 14 19 19 12 12 11 41 12
Karyotyp octomilky Drosophilla melanogaster Chromozómy jsou v párech, kromě pohlavních
Člověk Chromozómy v diploidních buňkách 2 n Celkem 46 (2 sady po 23) 22 párů autozómů (homologických chromozómů) a 1 pár gonozómů (pohlavních chromozómů) Samičí jsou homologické (XX) Samčí jsou nehomologické (XY)
Karyotyp - Člověk A A
Karyotypy - Larix Příbuzné druhy mohou mít podobné, i když poněkud odlišné chromozómy. Je možné studovat evoluci karyotypů
Lokus A lokus B lokus Od otce Od matky
DNA
Struktura DNA nosná kostra DNA homopolymerní řetězec cukr-fosfát Molekula DNA má tvar pravotočivé dvojšroubovice. Obě vlákna, skládající se z řetězce cukr (2 -deoxyribóza) a fosfát a dále z připojených nukleotidových bází, jsou vzájemně komplementárně spojena vodíkovými můstky mezi párovými bázemi, a sice adenin (A) s thyminem (T) a guanin (G) s cytosinem (C). A a T jsou spojeny dvěma vodíkovými můstky, zatímco C a G třemi. Na 1 otáčku připadá cca 10 nukleotidových párů. Haploidní genom člověka obsahuje cca 3, 000, 000 párů bází, které jsou rozděleny mezi 22 párů autozómů a 1 pár gonozómů.
Páry bází DNA Na 1 otáčku připadá cca 10 nukleotidových párů
Charakteristiky DNA Organizmus Escherichia coli Velikost genomu a 4, 7 Saccharomyces cerevisiae (kvasinka) Nicotina tabaccum 12 Počet genů Kódující DNA % 4000 100 6000 50 7800 Fritilaria davsii 295000 Picea abies 30000 35000 <3 Člověk 3000 50000 <2 a 106 párů bází u haploidní sady (čísla orientační)
Geny a Gen je možné definovat jako úsek DNA (sekvence nukleotidových bází, eventuelně tripletů), které po transkripci a translaci kódují syntézu specifického proteinu nebo RNA. Geny se skládají z: 1) kódujících sekvencí (nazývají se exony) 2) nekódujících sekvencí (introny) 3) regulační části (promotor, místo počátku transkripce) 4) koncových sekvencí Geny mohou mít různou délku (od cca 1000 až po několik set tisíc párů bází)
RNA v eukaryotních buňkách se vyskytuje RNA v několika formách: • mediátorová RNA • transferová RNA • ribozómová RNA
Proteosyntéza = exprese genů Proces, při kterém dochází k produkci proteinů na základě genetické informace uložené v DNA Je možné jej rozdělit do 2 dílčích částí: 1) Transkripce Nejprve je přepsána informace z DNA do m-RNA. Jedno vlákno dvoušroubovice DNA je při transkripci využito jako šablona, podle které RNA-polymerázy (syntetizující enzymy) vytvářejí m-RNA. Tato m-RNA následně přechází z jádra do cytoplazmy. Přitom prochází různými úpravami včetně vystřihání nekódujících sekvencí (intronů) a následného spojení exonů. Kódující m-RNA může být popsána jednotkami 3 nukleotidů, které se nazývají kodony (triplety).
DNA transkripce (přepis) matrice DNA do m-RNA vzniká primární transkript m -RNA, následně podléhá úpravám (zejména vystřihání nefunkčních intronů) přechází z jádra do cytoplazmy DNA
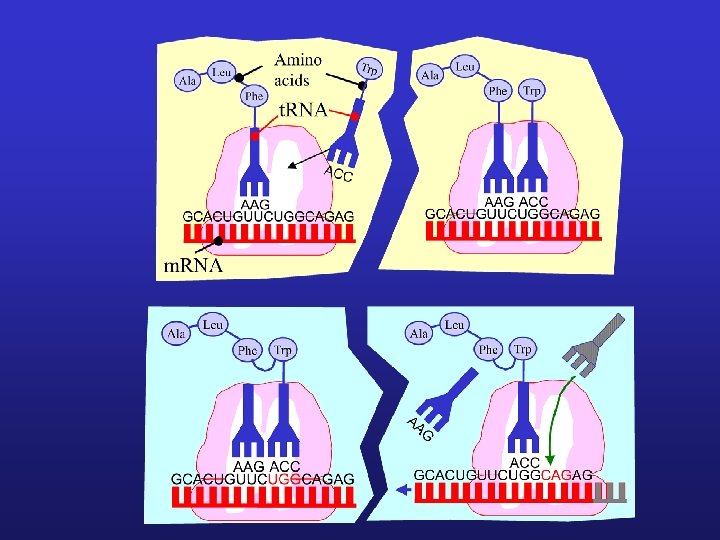
Kodony Kodon (triplet) = skupina tří sousedních nukleotidů na mediátorové (informační) RNA, které svým pořadím určují kvalitu AK a její polohu v peptidickém řetězci
t. RNA Transferové t. RNA jsou sestaveny z jednoduchého polynukleotidového řetězce, cca 80 nukleotidů Svým tvarem připomínají jetelový list Funkce t. RNA je přenos jednotlivých aminokyselin, z nichž se polypeptidy skládají
2) Translace = překlad proces překladu genetické informace z jazyka nukleotidů (kodonů) do pořadí aminokyselin v polypeptidovém řetězci Molekuly transferové t-RNA se svými antikodony (specifické sekvence 3 nukleotidů) vážou na komplementární kodony na m. RNA. Translace (syntéza proteinů) začíná v místě iniciačního kodónu AUG (tomu odpovídá t. RNA nesoucí AK methionin) a dále pokračuje ve smyslu kodónů AK se k sobě vážou za vzniku polypeptidu probíhá na ribozómech, které se přitom postupně pohybují podél řetězce m-RNA (po jednotlivých kodónech) Ukončení translace je zakódováno v kodónech UAA, UGA a UAG – po dosažení této sekvence dojde k uvolnění vytvořeného polypeptidu
Exprese genů = Proteosyntéza
Řetězec aminokyselin
Genetický kód DNA přenáší informaci do m-RNA ve formě kódu definovaného sekvencemi nukleotidových bází. - ribozómy se pohybují podél molekuly m-RNA a čtou její sekvence po 3 nukleotidech najednou (kodon). - kodón na m-RNA se páruje s komplementárním antikodónem na t-RNA - každá AK je specifikována určitým kodónem (nebo více kodóny) na m-RNA - RNA je konstruována ze 4 nukleotidů, existuje 64 možných kodónů (43) - tři z nich specifikují ukončení polypeptidového řetězce (nekódují AK), nazývají se STOP kodóny pak na 61 kodónů připadá 20 různých aminokyselin.
The Genetic Code 2 A = adenin G = guanin C = cytosin T = thymin U = uracil Ala: Alanin Cys: Cystein Asp: Asparagová kys. Glu: Glutamová kys. Phe: Fenylalanin Gly: Glycin His: Histidin Ile: Isoleucin Lys: Lysin Leu: Leucin Met: Methionin Asn: Asparagin Pro: Prolin Gln: Glutamin Arg: Arginin Ser: Serin Thr: Threonin Val: Valin Trp: Tryptofan Tyr: Tyrosin
Vlastnosti genetického kódu Tripletový Základní jednotkou je trojice nukleotidů – triplet, kodón, která při translaci kóduje 1 AK (nebo ukončení translace) Nepřekrývající se Záleží na začátku čtení (tj. od kterého nukleotidu se začne, neboť jinak by se změnil smysl informace), výjimka: viry Univerzální Smysl čtení tripletů platí pro veškeré živé organizmy Nadbytečný Většina z 20 AK je kódována více než 1 tripletem, u různých organizmů a různých typů NK může triplet kódovat odlišné kyseliny
Buněčný cyklus Cyklická reprodukce buněk od jednoho dělení ke druhému, zahrnuje 5 fází (včetně G 0 – klidové): G 1 -fáze: Nastává po dokončení předešlého dělení a vzniku dceřiných buněk Probíhá syntéza RNA a proteinů, buňka v této fázi roste S-fáze (= syntetická) Probíhá replikace DNA, chromozómy jsou „zdvojeny“ vzniklé sesterské chromatidy jsou spojeny v místě centromery + syntéza RNA a proteinů (zejména těch, které tvoří chromatin – histony) jako v G 1
G 2 -fáze Příprava pro vlastní dělení buňky pokračuje růst, intenzívně se tvoří RNA a proteiny M-fáze Vlastní dělení buňky na 2 geneticky identické dceřiné buňky Zahrnuje 4 fáze: Ø Ø Profáze Metafáze Anafáze Telofáze
Mitóza aaaaaaaaaaaaa replikace DNA (S-fáze) homologické chromozómy sesterské chromatidy po replikaci aaaaaaa segregace chromozómů sesterské chromatidy se oddělí
Profáze Ø nastává kondenzace chromozómů Ø mizí jaderná membrána a jadérko Ø začíná se tvořit mitotické vřeténko Ø u pólů buňky se morfologicky zvýrazňuje část centrozómu v podobě 2 centriol
Metafáze Ø chromozómy jsou maximálně nahuštěné a uspořádané v rovníkové rovině Ø sesterské chromatidy jsou zřetelně oddělené, ale stále spojené v centromeře
Anafáze chromatidy (nyní již nové chromozómy) po rozdělení centromery putují k pólům pohyb je umožněn zkracováním fibril dělícího vřeténka
Telofáze Ø shlukování chromozómů u pólů Ø dekondenzace Ø tvorba jaderné membrány V rovníkové rovině se objevují mikrofibrily odškrcující nové buňky
Meióza aaaaaa Proces crossing over A homologické chromozómy sesterské chromatidyi AAAAA Oddělení homologických chromozómů I sesterské chromatidy nadále spojeny centromerou AAAAaaaaa Oddělení homologických chromozómů II Gamety
Meióza Proces dělení jádra, při kterém se tvoří haploidní gamety Dělí se na 2 základní etapy: 1) Heterotypické dělení (= redukční, meióza I) Profáze I – časově zaujímá 90% ØLeptotene chromozómy tvořící klubko začínají kondenzovat ØZyotene homologické chromozómy se párují, vytvářejí bivalenty ØPachytene nesesterské chromatidy spárovaných chromozómů se překřižují (= crossing over), tj. chromozómy si vymění části chromatid (= rekombinace) ØDiplotene, diakineze
Crossing over
Metafáze I chromozómy (zde páry nerozdělených bivalentů) se řadí v rovníkové rovině Anafáze I homologické chromozómy s vyměněnými nesesterskými chromatidami se NAHODILE rozchází k pólům (centriolám) Telofáze I vznikají 2 dceřiné buňky = gametofyty 2. řádu
2) Homeotypické dělení (meióza II) do značné míry podobné mitóze v anafázi II se sesterské chromatidy s rekombinovanými segmenty oddělí v telofázi II vznikají z gametofytů 2. řádu 2 dceřiné buňky = gamety (n) Výsledkem celé meiózy je VZNIK 4 HAPLOIDNÍCH GAMET z původní jedné zárodečné buňky
GENETICKÉ DŮSLEDKY MEIÓZY Ø počet možných kombinací chromozómů ve vzniklých gametách je 2 n (u rodu Pinus 4096) Ø kromě nahodilých kombinací navíc crossing over u nesesterských chromatid Výsledkem současného působení obou procesů je existence prakticky nekonečné genetické variability (žádní 2 jedinci nejsou u pohlavního rozmnožování identičtí – kromě monozygotických dvojčat)
Rodiče potomstsvo přesně polovina genetické výbavy od každého z rodičů průměrně jedna čtvrtina genetické výbavy od prarodičů, množství se může lišit
Oplodnění haploidní gamety diploidní zygota
Dědičnost na úrovni organizmu Zakladatel moderní nauky o dědičnosti byl Johann Gregor Mendel: Ø nejvhodnějším přístupem ke studiu dědičnosti a proměnlivosti znaků je analýza hybridologických experimentů (křížení) Ø pro studium základních genetických zákonitostí nejprve dědičnost kvalitativních znaků
Křížení = hybridizace vzájemné oplozování organizmů s různými genotypy, tj. generativní spojení 2 různých gamet písmenem P se označuje rodičovská generace (parentes) písmenem F (filius, filia) + indexy generace potomků vzniklých křížením (1. , 2. , 3. , …. filiální generace) např. P: AA x aa F 1: Aa F 2: AA : 2 Aa : aa
Podvojné založení dědičnosti Znak je u diploida založen 2 formami genu – alelami (od otce a od matky), které jsou uloženy na příslušných lokusech homologních chromozómů, mohou být dominantní a recesivní Výjimka: dědičnost znaků vázaných na pohlaví A A lokus a Alely mohou být na homologických chromozómech: stejné kvality – homozygot v daném znaku B lokus různé kvality – heterozygot v daném znaku
Při studiu dědičnosti Mendel použil hrách setý (Pisum sativum) Ø je samosprašný Dominantní homozygot Ø sledoval pouze alternativní znaky Ø křížil čisté linie, tj. homozygoty v daných znacích (7 znaků, např. tvar semen, barva květů) P: AA x aa F 1: Aa F 2: AA : 2 Aa : aa Recesivní homozygot
dominantní homozygot x recesivní homozygot P: AA x aa A A a Aa Aa recesivní homozygot x heterozygot P: aa x Aa A a a Aa aa heterozygot x heterozygot P: Aa x Aa A a A AA Aa aa dominantní homozygot x heterozygot P: AA x Aa A A A a AA Aa
Dihybridní křížení ve 2 znacích zároveň A žlutá semena a zelená semena P: AABB x aabb B kulatá semena b hranatá semena F 1: Aa. Bb F 2: ♀♂ AB Ab a. B ab AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb Aabb aa. Bb aabb
Dihybrid tvoří 16 zygotických kombinací, u kterých můžeme zjistit Ø 9 odlišných genotypů Ø 4 odlišné fenotypy
Mendelovy zákony Byly formulovány na základě uvedených pokusů Ø Pravidlo o uniformitě hybridů F 1 generace a identitě reciprokých křížení Ø Pravidlo o zákonitém štěpení v potomstvu hybridů F 2 – princip segregace Ø Pravidlo o volné (nezávislé) kombinovatelnosti vloh Ø Pravidlo o čistotě gamet
Podmínky platnosti Mendelových zákonů ØJen pro monogenní znaky ØJen pro úplnou dominanci alel ØPři úplné náhodnosti křížení ØPříslušné geny musí sídlit na autozómech, event. na homologických úsecích gonozómů
VZTAHY MEZI ALELAMI 1 GENU Úplná dominance dominantní alela svým účinkem převládá a stačí k úplnému projevu znaku (AA i Aa mají stejný fenotypový projev > aa) Neúplná dominance dominantní alela u heterozygota plně nepřekrývá svým účinkem alelu recesivní, na fenotypu se projeví obě nestejnou intenzitou AA > Aa > aa
Kodominance u heterozygota dominují obě alely společně, jejich produkty jsou na sobě nezávislé (ve fenotypu se projeví obě alely plným projevem) Superdominance znak u heterozygota je vyjádřen silněji než u obou homozygotů toto může být mnohdy výhodné, a sice díky přítomnosti obou alel AA < Aa > aa
GENOVÉ INTERAKCE jev, kdy 1 znak vzniká spolupůsobením 2 a více genů, a to umístěných na jednom chromozómu nebo na různých důsledkem genových interakcí je snížení počtu fenotypových tříd (dihybrid 9 : 3 : 1, zde např. pouze 15 : 1) Komplementární účinek patří mezi nejčastější interalelické interakce, k projevu znaku dojde tehdy, je-li přítomna alespoň 1 dominantní alela z každého z obou genů (u dihybrida 9 : 7)
Epistáze jev nadřazenosti jednoho genového páru nad druhým, jedná se o analogii dominance a recesivity v rámci 2 a více genů typickým znakem u rostlin i zvířat je zbarvení Ø dominantní alela A jednoho lokusu je nadřazena nad dominantní alelou druhého lokusu B, tzn. existuje-li alespoň 1 alela A (tedy u AA nebo Aa), pak dominantní alela D se NEMŮŽE PROJEVIT (12 : 3 : 1)
Pleiotropie jev, kdy se 1 gen (1 pár alel) podílí svým účinkem na realizaci několika znaků (např. opakované působení genu během ontogeneze)