Vstupy centrlnch regultor checkpoints reakce na pokozen a







 UV wt rad 9 (mrtvé se")
 wt,")
 • Existují mechanismy, které průběžně monitorují úplnost a „zdraví“ důležitých")

")
")

")


")
")
")






 Proč jsou Hartwellovi mutanti")


 Y 15 T 14")
")









- Slides: 41
Vstupy centrálních regulátorů: checkpoints – reakce na poškození a zpětná vazba
„cell cycle engine“ vstupy velikost signály poškození. . . výstupy gen. exprese morfogeneze. . .
? ? ? • Why do events occur in a particular order? • How are superficially unrelated events kept in phase? • How are growth and division co-ordinated? • How does the cell know where to locate new structures? (A. E. Wheals, 1976)
L. Hartwell a T. Weinert, 80. 90. léta cdc 9
Ale proč? ?
K čemu je dobrá ligáza?
Je zástava cyklu důsledkem zlomů DNA?
Mutace zvyšující citlivost ke zlomům: rad (radiation sensitive) UV wt rad 9 (mrtvé se barví methylenovou modří)
UV dělá zlomy. . . a co defekt ligázy? ? (občas ztráta chromosomu) wt, 37 o. C rad 9, 25 i 37 o. C RAD 9 nutný k bloku cyklu při poškození! cdc 9, 37 o. C cdc 9 rad 9, 37 o. C (Hartwell and Weinert 1988)
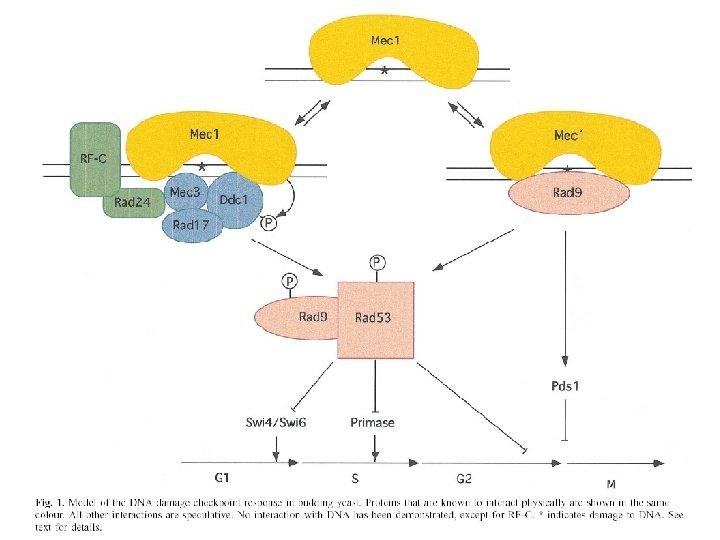
Model „kontrolních bodů“ (checkpoints) • Existují mechanismy, které průběžně monitorují úplnost a „zdraví“ důležitých mechanismů BC a v případě poruchy cyklus zastaví, aby byl čas na opravu. • RAD 9 je součástí dráhy, která takto monitoruje celistvost DNA (DNA damage checkpoint). poškozená DNA v S nebo G 2/M G 2 M
Jak funguje RAD 9 checkpoint? • 2 epistatické třídy RAD genů – RAD 17, RAD 24, MEC 3 a DDC 1 – RAD 9 • protein kináza MEC 1 – detektor?
(Melo and Toczyski 2002)
Cíl 1: transkripce cyklinů (CLN via SCB, CLB via MCB)
Cíl 2: replikace - přes primázu
Cíl 3: securin (inhibitor separace chromosomů, substrát APC, který vyvazuje separasu)
Tenodera aridifolia sinensis Replikace není jediným zdrojem rizika. . .
Chromosomové určení pohlaví u kudlanky ♀: X 1 X 2 ♂: X 1 X 2 Y
Anafázi lze obnovit mechanickým tahem za kinetochor! Anafáze Blok Anafáze (Li a Nicklas 1995)
Jak kinetochor ví, že se za něj tahá? (anti-phosphoprotein Ab)
(Li and Nicklas 1997)
Jak je to u kvasinek? • benomyl – destrukce vřeténka a M arrest • MAD/BUB geny: mitotic arrest deficient/budding uninhibited by benomyl wt, benomyl mad/bub, benomyl
„Spindle checkpoint“ • Aktivace u kvasinek: – benomyl, nocodazol a jiné mt jedy – mutace SPB – mutace centromer (CDEIII) – nadpočetné centromery (mnoho různých YCp) • Aktivace u hmyzu: – nepřichycené chromosomy
Blok anafáze: via APC
Co když se chromosom ztratí později? Bub 2: část „dvousložkového“ GAP (připomínka: Cdc 14 je fosfatáza nezbytná pro mitotic exit)
Regulace přechodu Ana/Telo
Spindle checkpoint - cíle securin/separáza telofázní regulátor APC transkripce CKI
A to ještě není vše! cdc 24 cdc 42 (1995) Proč jsou Hartwellovi mutanti dvojjaderní, ale v jiném genetickém backgroundu jen 1 G 2 jádro?
iniciace terminace segregace Absence pupene zdržuje mitosu!
Obecně: cell morphogenesis checkpoint Senzor: proteinové komplexy asociované se septiny Y 19 F a někt. kmeny standardní wt Cíl: inhibiční fosforylace CDC 28 (Y 19)!
Pro kontrolu pupenem nutná fosforylace Y 19 (homolog Y 15) Y 15 T 14 Y 15 T 161 Y 15 T 14 T 161 Y 15 cdc 25 T 14 T 161 wee 1 Y 15 T 161
Cíle – širší pohled (Saccharomyces wee 1)
? ? ? • Why do events occur in a particular order? • How are superficially unrelated events kept in phase? • How are growth and division co-ordinated? • How does the cell know where to locate new structures? (A. E. Wheals, 1976)
Kontrola velikostí: necyklující cyklin CLN 3 velikost CLN 3 START SWI 4/6 CLN 1, 2
Jaký to má vztah k živinám? A jak je kvasinka vnímá? • „Start II“ mutace: – blok ve Startu bez růstu – „kvasinka si myslí, že hladoví“ • CDC 25: GEF pro RAS!
CDC 25 stimuluje produkci c. AMP GEF adenylátcykláza Cdc 25 Ras-GDP Ras-GTP Cyr 1 fosfodiesteráza Pde Ira 1, 2 GAP ATP AMP inhibice nebo stimulace Startu?
Regulace proteinkinázy A pomocí c. AMP adenylátcykláza Cyr 1 (Cyr 1: sporulace na bohatých médiích etc. ) AMP ATP proteinkináza A (PK-A) Tpk 1, 2, 3 1 y c B (zase jedna kináza regulovaná inhibitorem) Tpk 1, 2, 3 ? 1 y c B (Bypass of Cyr)
Kvasinková RAS/c. AMP dráha GEF adenylátcykláza Cdc 25 Cyr 1 Ras-GDP fosfodiesteráza Ras-GTP Ira 1, 2 GAP Pde AMP ATP proteinkináza A (PK-A) Tpk 1 y c B inhibice nebo stimulace Startu?
asynchronní kultura Předpokládali bychom, že c. AMP stimuluje, ale. . . (M. D. Baroni, Milano, Italy – Nature 371: 339, 1994
a-factor release . . . c. AMP inhibuje BC, aby mohla vzrůst kritická velikost!
c. AMP zprostředkuje spřažení cyklu a růstu u S. cerevisiae • Cíl regulace přes Tpk/PK-A: nastavení citlivosti transkripce CLN 1 a CLN 2 vůči hladině CLN 3 • Čím víc cukru, tím víc c. AMP a tím víc CLN 3 = větší velikost před startem! • c. AMP podobně reguluje i anafázi a mitotic exit (srv. S. pombe!) – inhibice APC prostřednictvím PK-A