Virus Attachment Entry and Uncoating Virus Properties Virus






 Adsorption (attachment): • random collision • interaction between specific proteins on")
 Adsorption (attachment): – some viruses may use more than one host")
 Entry (penetration): • 2 mechanisms - endocytosis - fusion of virus")

 that the virus is able to")

 in triangular face")
 phage structure From The Biology of Viruses, Voyles, Mc. Graw Hill,")



 - a virus with")

")

")






 has a")



 • • • A classic example of a receptor/co-receptor requirement A specific")
 - I • HIV enters by a quite different route")
 initially")
 - II • • If p. H is not required")

 • Classically all retroviruses were")
 • Virus receptor is a lipid (phosphatidyl serine;")


 •")





 • Picornaviruses bind to a variety of specific cell surface molecules -")
 • Although not p. H dependent, poliovirus")




 • VP 4")






 is shed phosphorylation dependent •")
 contains NLSs and the RNPs are")
 can only replicate in")
- Slides: 65
Virus Attachment, Entry and Uncoating
Virus Properties • Virus is defined as a nucleoprotein complex which infects cells and uses their metabolic processes to replicate • Smallest known infective agents • Metabolically inert - no metabolic activity outside host cell; must enter host cell to replicate • Most are highly species specific
Virus versus Virion • Virus is a broad general term for any aspect of the infectious agent and includes: • the infectious or inactivated virus particle • viral nucleic acid and protein in the infected cell • Virion is the physical particle in the extracellular phase which is able to spread to new host cells; complete intact virus particle
General points - virus entry • • The first event in any virus life-cycle - often limits infection to the “correct” cell Can be primary determinant of tropism – – • • Tissue tropism - e. g. measles (skin cells) vs. mumps (salivary gland) Species tropism - e. g. togavirus (both insect/mammalian cells), poliovirus (primate cells), T 4 phage - (few strains of E. coli) Binding - initially electrostatic, based on charge ± p. H, specific ions - followed by local hydrophobic interactions Initial binding is often low affinity, but high avidity (tight binding) due to multiple binding sites The virus binds to a receptor on the cell surface - can be ubiquitous/specific, with variable density Initial binding is followed by penetration and subsequent uncoating
General points II - virus entry • Whether or not the virus is enveloped makes a big difference - at least for penetration • All viruses must cross a lipid bilayer, plant and bacterial viruses must also cross a cell wall • Uncoating means that the stable virus stucture must become unstable – -transition from extracellular (chemical) form to intracellular (biological) form – There must be some sort of “trigger” or regulated disassembly process
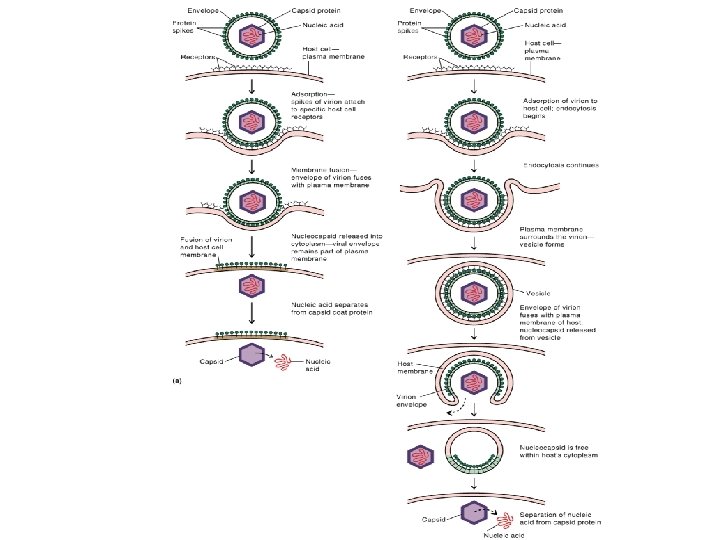
Overview Name Process 1 2 3 Attachment Entry Uncoating Virus attach to cell membrane By endocytosis or fusion By viral or host enzymes, separates NA from protein coat 4 5 6 7 8 Biosynthesis Maturation Release Shedding Infecting new host Production of NA and proteins NA and capsid proteins assemble By budding (enveloped viruses) or rupture 6
Viral Replication i) Adsorption (attachment): • random collision • interaction between specific proteins on viral surface and specific receptors on target cell membrane (tropism) • not all cells carrying a receptor for a particular virus can be productively infected by that virus
Viral Replication i) Adsorption (attachment): – some viruses may use more than one host cell receptor (e. g. HIV) – able to infect a limited spectrum of cell types (host range) – most neutralizing antibodies are specific for virion attachment proteins
Viral Replication ii) Entry (penetration): • 2 mechanisms - endocytosis - fusion of virus envelope with cell membrane iii) Uncoating: • release of viral genome • cell enzymes (lysosomes) strip off the virus protein coat • virion can no longer be detected; known as the “eclipse period”
Attachment: Cell Receptor • Virus may bind up to three different cell receptors in succession: – Low affinity receptor - in high abundance, virus contacts cell surface – Primary receptor - in lower concentration – Co-receptor – follows binding of primary receptor
Attachment: Specificity • Host Range - the organism(s) that the virus is able to infect (narrow or wide) i. e. plant, animal, human • Tissue Tropism- the cell type(s) a virus is able to infect i. e. skin, oral, GI, CNS
Attachment: Binding • 3 -D fit between viral ligand cell receptor • Mainly weak electrostatic charges. • Evidence for this is interaction may require: – specific p. H – specific ionic strength – presence of specific ions i. e. Ca++, Mg++
Attachment: Nonenveloped Picornavirus • Virus ligand - a deep cleft (“canyon”) in triangular face of capsid (viral proteins VP 1, VP 2, VP 3) • Binds to cell receptor ICAM – 1 (intracellular adhesion molecule 1), normal function is to bind cells i. e. WBC
T-even (T 2) phage structure From The Biology of Viruses, Voyles, Mc. Graw Hill,
Entry of T-even bacteriophage - binding • Best understood = T 4 phage (the virus in the Hershey-Chase experiment) • Initial binding is reversible and electrostatic - the outer-most part of the long tail fiber binds to surface lipopolysaccharides (LPS) of the bacterium (binding can occur in vitro, and is competed by specific sugars) - a non-specific receptor • Binding is “additive” until all six tail fibers are bound • Binding of 3 fibers is needed to initiate infection From Introduction to Modern Virology, Dimmock & Primrose, Blackwell The virus may “browse” the surface - looking for a suitable site for penetration (possibly sites of cell wall synthesis where the outer and plasma membranes are close together). Note - this is a multi-valent interaction
Entry of T-even bacteriophage - binding II • The receptor binding sites of the short tail fibers are now exposed and bind (also to LPS)- now binding is essentially irreversible • Conformational change in the baseplate hexagon to extended star-shaped conformation • Initiates sheath contraction ( to 37% of its original length)
Entry of T-even bacteriophage - penetration • Often referred to as a “hypodermic syringe” • Sheath of the helical tail slips and forms a shorter helix. • The tube itself stays the same with the end result that the tube is pushed down and contacts the membrane - note the tail does not directly punch through • Lysozyme molecules are releases which forms a pore through which DNA enters From The Biology of Viruses, Voyles, Mc. Graw Hill, DNA is under considerable pressure and seems to exit automatically once the base palte is opened up Other phage, e. g. T 3, have a motor protein to reel out the DNA
Entry of other ‘phage - I • Phage λ (DNA) - a virus with a longer, but simpler tail than T 4 From The Bacteriophages vol 1, ed Calender, R. , Plenum Press the single tail fiber (J protein) binds to lam. B (the maltose transporter) - an example of a specific receptor lam. B is inducible. This means the virus only infects in the presence of high nutrients. Also needs Mg 2+ - binding is electrostatic - an example of tropism Penetration requires the bacterial pts protein (also part of the maltose transporter) - the co-receptor attachment and penetration require different viral proteins
Entry of other ‘phage - II • PRD 1 - an icosahedral phage with an internal membrane • For gram -ve bacteria (with two layers of lipid separated by peptidoglycan) phage entry is a challenge – – – 1) Binding 2) Conformational change -> dissociation and opening of 14 nm hole in the capsid 3) Second conformational change converts internal envelope to tubule, which delivers the DNA Phage encodes 2 proteins (P 5 and P 17) that have peptidoglycan-hydrolyzing activity - equivalent protein in T-even phage = gp 5 (lysozyme activity) From Rydman and Bamford (2002) ASM News 68 330
Entry of other ‘phage - III • Enveloped RNA phage - φ6 (Phi 6) • These phage bind to pili, the pilus then retracts down to the outer membrane, the virus undergo fusion, enzymatically destroys the peptidoglycan cell wall (p 5 protein) and then penetrates the plasma membrane From The Bacteriophages vol 2, ed Calender, R. , Plenum Press The Hershey-Chase experiment is now no longer valid, as (most of) the protein (35 S) has entered the cell along with the nucleic acid (32 P).
Plant viruses • Plants have a thick, rigid cell wall • Generally plant viruses do not have specific entry mechanisms, but rely on – A) introduction into the cell by a vector (insect) - most common – B) mechanical injury – C) direct cell-cell transmission (via plasmadesmata and viral movement protein) • This is fine if you are a non-enveloped virus, but enveloped plant viruses do exist (bunya-, rhabdofamilies – These viruses must fuse their envelope
Binding of animal viruses • Occurs via receptors on the cell surface (plasma membrane) Protein (glycoprotein) Carbohydrate Lipid (glycolipid) From Principles of Virology, Flint et al. ASM Press Receptor utilization plays a major role in virus tropism / pathogenesis
Entry / Uncoating • Entry is the mechanism used by the virus to penetrate into the host cell • Uncoating is the separation of the nucleic acid from the capsid, and refers to changes that occur to make the viral nucleic acid ready for expression
Principles of virus penetration • Viruses can penetrate directly at the plasma membrane, or via endosomes
Penetration of Enveloped Animal Viruses • • • Envelope = fusion Semliki Forest virus (SFV) a togavirus - the classic virus for entry studies Early experiments (early 1980 s) by electron microscopy showed entry into vesicles - now known to be clathrin-coated (clathrin-coated vesicles, or CCVs) – The virus then enters the endosome (“early” endosome) Figure courtesy of A. Helenius The very high particle: pfu ratio (approaching 1: 1) of SFV ensures that all the virus particles are part of the “real” entry pathway
Endosomes and virus entry • • Endosomes are used by cells for nutrient and growth factor uptake The virus “hijacks” the cellular pathway One key feature of endosomes is their progressive acidification due the action of the vacuolar H+/v. ATPase Endosomes do much more than provide low p. H – – – Deliver through cortical actin and microtubule-mediated transport in the cytosol Specific redox/ionic environment Defined lipids for fusion/penetration From Cell Biology, Pollard and Earnshaw, Saunders
• The lowered p. H causes conformational changes in the spike glycoprotein, and the exposure of a fusion peptide • This is the “trigger” needed for virus entry • In most cases a p. H of around 6. 2 -6. 5 is sufficient for fusion - fusion occurs in the early endosome • Entry and infectivity (in cell culture) can be blocked by : – – 1) addition of a weak base (e. g. NH 4 Cl) that neutralize the endosome 2) drugs that target the v. H+/ATPase (e. g bafilomycin A) 3) drugs that break down the proton gradient (e. g. monensin) 4) exposure of the virus to a low external p. H Fusion can be induced at the cell surface by exposure to low p. H
Influenza virus binding - I • • • Binds to cell surface carbohydrate - sialic acid Ubiquitous/non-specific receptor In principle, this can be present as part of glycoprotein or glycolipid Specific requirement for α 23 and α 2 -6 linkages - gives different tropism for avian vs. human cells (pigs have both) From Principles of Virology, Flint et al. ASM Press The first virus receptor to be identified - principally due to the fact that there is a receptor-destroying enzyme associated with the virus
Influenza virus binding - II • The major influenza glycoprotein, hemagglutinin (HA) has a specific sialic acid-binding site on its “top domain” - From Principles of Virology, Flint et al. ASM Press HA mediates both binding and penetration
Penetration of influenza virus • • Influenza virus requires a lower p. H (5. 0 -5. 5) and enters the “late” endosome, but fusion occurs before entry into the lysosome (this avoids degradation) The acid-triggered fusion event is well understood - a conformational tail forms a rigid “six-helix bundle” or “coiled-coil” of α-helices, which flips the fusion peptide out and allows insertion into the membrane – Note the fusion peptide is “external” The “trigger” is irreversible - this means that exposure of virions to low extracellular p. H will destroy infectivity From Principles of Virology, Flint et al, ASM Press
From Principles of Virology, Flint et al, ASM Press • The low p. H has a second very important role for influenza entry - the virus contains an ion channel in its envelope (M 2). • The presence of M 2 allows acidification of the virus interior, and promotes uncoating of the M 1/v. RNPs • Drugs that block M 2 block infection - amantadine. This is highly specific for the viral M 2 ion channel, with no effect on the cellular H+/v. ATPase
Fusion of an enveloped virus Model for viral membrane fusion mediated by class I fusion roteins. Influenza virus, which is internalized into an endosome, is shown as an example. In the native state of the fusion protein — which is a trimer — most of the surface subunit (green) is exposed. Part of the transmembrane subunit, including the fusion peptide, is not exposed. Following fusionactivating conditions, conformational changes occur to 'free' the fusion peptide (red) from its previously unexposed location. In the case of influenza HA, this occurs by a 'spring-loaded' mechanism. The 'pre-hairpin' intermediate spans two membranes — with the transmembrane domain positioned in the viral membrane and the fusion peptide inserted into the host-cell membrane. The pre-hairpin intermediate forms a trimer of hairpins, and membrane fusion occurs, which leads to pore formation and release of the viral genome into the cytoplasm. From Dimitrov (2004) Nature Reviews Microbiology 2: 109 -122
Retrovirus (HIV) • • • A classic example of a receptor/co-receptor requirement A specific receptor Binds initially to CD 4 - present on immune system cells (T-cells) - gives the virus tropism for the immune system This is not enough - the virus also binds to a chemokine co-receptor (eg CCR 5, CXCR 4) present on a sub-set of cells (macrophages / T-cells) Gives even more precise tropism The virus binds to both receptors via the gp 120 glycoprotein
Penetration of retrovirus (HIV) - I • HIV enters by a quite different route • Entry is not low p. H-dependent (no inhibition by NH 4 Cl etc), and fusion occurs directly with the plasma membrane From Principles of Virology, Flint et al, ASM Press
Attachment: Enveloped HIV Virus • Host cell protein in virus envelope (cyclophilin A) initially binds HIV to low affinity receptor (heparin sulfate) of the cell • Followed by binding of viral ligand (gp 120) to primary receptor (CD 4) on T helper cells, macrophages, and glial cells • Binding of gp 120 to CD 4 results in conformational change of gp 120, which then binds to chemokine coreceptor CXCR 4 on T lymphocytes or CCR 5 on macrophages
Penetration of retrovirus (HIV) - II • • If p. H is not required for fusion, what is the trigger ? ? Following receptor binding a conformational change (also the formation of a coiled coil) occurs in the HIV-1 gp 120 molecule exposes its fusion peptide (present on gp 41 - the second half of the gp 160 Env protein) From Principles of Virology, Flint et al, ASM Press HIV has a receptor (CD 4) and a coreceptor (CCR 5 or CXCR 4)
• HIV virions are able to gain access to their host cells by way of viral host-cell membrane fusion (Gallo et al. , 2003). • The viral envelope gp 120 first recognizes its primary receptor on host cells, CD 4 (Dalgleish et al. , 1984; Ugolini et al. , 1999), using a binding motif contained within its second constant (C 2) region (Kwong et al. , 1998). • This interaction gives rise to a conformational change in gp 120 which exposes its third variable loop (V 3) which contains a consensus amino acid motif that allows for binding to a seven transmembrane-spanning chemokine co-receptor (Jones et al. , 1998; Kwong et al. , 1998; Berger et al. , 1999). • Both interactions are necessary for viral fusion and entry (Sattentau and Moore, 1991).
Entry of avian leukosis virus (a model, simple, retrovirus) • Classically all retroviruses were thought to be p. H-independent • More recently ALV has been proposed to require low p. H, but in addition to its receptor-induced conformational change • Entry is occurring via endosomes in this case
Entry of vesicular stomatitis virus (VSV) • Virus receptor is a lipid (phosphatidyl serine; PS) – a unique example ? ? • Very wide infection range (all cells have PS) one of the most promiscuous viruses out there • Fusion etc is similar to influenza…. . – Both VSV G and influenza HA are referred to as type I fusion proteins • with two main differences – The trigger is reversible – The p. H threshold is less stringent (approx. p. H 6. 5). Fusion is though to occur from the “early” endosome
Type I and type II fusion proteins • Type I is the most common and understood fusion protein – Influenza, VSV, retrovirus • Type II fusion proteins are not proteolytically activated, have internal fusion peptides and no “coiled-coil” form; they are principally β-sheet • Flavivirus (TBE), and Alphavirus (SFV)
SFV and TBE - alternative ways to expose fusion peptides • In SFV the fusion peptide is protected by E 2 • In TBE the flat E protein rotates and twists
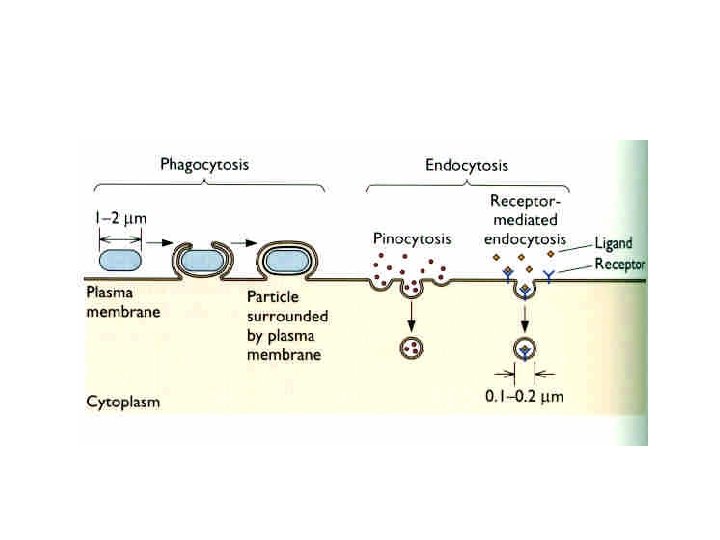
Entry: Nonenveloped Virus • Receptor-mediated endocytosis • Clathrin coated pits (seen by EM) • Invagination and pinching off of the membrane • Forms an intracellular endosome containing the virus • Endosome becomes acidified
Uncoating: Nonenveloped Virus • Low p. H causes conformational changes in capsid protein (hydrophobic region interacts with membrane forming a pore) • Viral nucleic acid released
Clathrin vs. non-clathrin internalization • Most viruses were originally assumed to use clathrin as a route into the cell • Used by SFV, VSV, adenovirus etc • Other routes of entry exist and can be used • Caveolae (as used by SV 40) are the best characterized) • Influenza and rotavirus are other examples • In most cases non-clathrin pathways are illdefined
Dynamin is a GTPase that acts to “sever” the necks of the endocytic vesicle It is not specific to clathrincoated vesicles Dominant-negative mutant (K 44 A) inhibits endocytosis Eps 15 binds to AP-2, the clathrin adaptor protein It is specific to clathrincoated vesicles Dominant-negative mutant (Eps 15 delta 95 -295) inhibits endocytosis From Biochem. J. (2004) Immediate Publication, doi: 10. 1042/BJ 20040913 Cargo- and compartment-selective endocytic scaffold proteins Iwona Szymkiewicz, Oleg Shupliakov and Ivan Dikic
Lipid rafts • Detergent-resistant domains in cell membranes • Enriched in cholesterol and sphingomyelin Play a very important role in virus budding Can also be important for virus entry , esp non-clathrin endocytosis e. g SV 40 From Munro S. Cell. 2003 Nov 14; 115(4): 377 -88. Lipid rafts: elusive or illusive?
Herpesviruses • • • A complex system Herpesviruses have 10 -12 surface glycoproteins Binds initially to heparan sulfate (via g. C) – used by a multitude of different viruses - non-specific – An attachment or “capture” receptor • Subsequently binds to a co-receptors that allows entry (via g. D) - herpesvirus entry mediator - specific – A fusion receptor – – Hve. A Hve. B Hve. C Hve. D TNF-R Nectin 2 (Prr 2) Nectin 1 (Prr 1) PVR • Different herpesviruses use different receptors • But very different viruses can use the same receptor – e. g. pseudorabies virus and polio virus – Another example = CAR - the coxsackie/adenovirus receptor
Poliovirus/Rhinovirus (Picornaviridae) • Picornaviruses bind to a variety of specific cell surface molecules - these are specific proteins – Binding occurs via canyons (depressions) in the virus surface From Principles of Virology, Flint et al. ASM Press Similar viruses can have quite distinct receptors
Penetration of non-enveloped viruses • Rhinovirus/Poliovirus (Picornavirus) • Although not p. H dependent, poliovirus may still enter through the endosome • Interaction of poliovirus with PVR causes major conformational changes in the virus leads to the formation of the A particle physically swollen (less dense) From Principles of Virology, Flint et al, ASM Press
• A particles are now hydrophobic. Viruses have apparently lost VP 4, and the hydrophobic core is exposed on the virus surface Penetration might be controlled by sphingosine, a lipid present in the “pocket” -- or (more likely) by the pocket allowing “breathing” of the capsid • With a non-enveloped virus, fusion is not possible. Instead picornaviruses form a membrane pore From Principles of Virology, Flint et al, ASM Press Parvoviruses may contain a phospholipase A 2 activity in their capsid protein The specific lipid composition of endosomes may be crucial for some viruses
Adenovirus • A relatively complex system • Receptor and co-receptor • Clathrin-mediated • • • endocytosis Instead of forming a discrete pore, adenovirus ruptures or lyses the endosomal membrane The trigger is low p. H, via the penton base protein The virus undergoes proteolytic cleavage - by virus-encoded proteases
SV 40 • Entry occurs via endocytosis but in a clathrin-independent manner • Entry does not depend on low p. H • The virus enters through “caveolae” - a specialized endocytic vesicle that forms upon specific cellular signaling induced by virus binding • Receptor is combination of a protein (MHCI) and a glycolipid (sialic acid)? • The “caveosome” containing the virus is delivered to the ER Caveolae are specialized lipid rafts
Reovirus • The rare example of a virus requiring the lysosome • Reoviruses have a complex double capsid, which is very stable to low p. H (gastro-intestinal viruses; rotavirus) • The lysosomal proteases degrade the outer capsid to form a subviral particle i. e degradation by cellular proteases From Principles of Virology, Flint et al, ASM Press
Rotavirus entry • Trypsin cleavage of VP 4 (= spike protein) • VP 4 becomes VP 8* and VP 5* • Transient exposure of hydrophobic peptide • Trimeric coiled coil formation From Dormitzer et al (2004) Nature 430: 1053 Comparable to influenza HA ?
The problem of cytoplasmic transport • • • Assume the virus in question has undergone receptor binding and penetration - ie the virus/capsid in the cytoplasm. The cytoplasm is viscous and the nucleus is often a long distance from the site of entry. This is especially true for specialized cells such as neurons μm μm 1 cm polio 61 yr HSV 231 yr Table box 5. 2 From Sodeik, Trends Microbiol 8: 465
Microtubules and virus entry VSV/Rabies, influenza Adenovirus From Sodeik, Trends Microbiol 8: 465 Herpesvirus • To facilitate transport viruses often bind to the cytoskeleton and use microtubule-mediated motor proteins for transport, i. e. dynein
Nuclear Import • Why replicate in the nucleus? What are the “benefits? ” • • • DNA viruses - need cellular DNA polymerase and/or accessory proteins (eg topoisomerase) All DNA viruses replicate in the nucleus exception = Pox viruses (even these will not replicate in an enucleated cells or cytoplast) Almost all RNA viruses replicate in the cytoplasm, and most will replicate in a cytoplast Principal exceptions = retroviruses (these have a DNA intermediate and influenza virus (has a spliced genome)
What are the “problems” with nuclear replication? • • • An additional barrier during genome transport The nucleus of a eukaryotic cell is surrounded by a double lipid bilayer - the nuclear envelope. The nuclear envelope is studded with transport channels the nuclear pores From Flint et al Principles of Virology ASM Press
Parvovirus • • • Possibly the simplest example of nuclear entry Small icosahedral DNA virus (18 -26 nm diameter) Enters through endosomes (p. H-dependent) VP 1 contains a nuclear localization signal (NLS) From Flint et al Principles of Virology ASM Press Basic amino acids The NLS binds to cellular receptors (karyopherins or importins) that carry proteins into the nucleus But, the NLS is hidden on the inside of the capsid Therefore a conformational change must occur to expose the NLS
Adenovirus • • • Contains NLSs on its capsids, binds microtubules But, The functional size limit of the nuclear pore is 26 nm The virus is therefore transported as far as the pore. It docks to the nuclear pore and then undergoes final disassembly, and the DNA is “injected” into the nucleus with DNA binding proteins attached Specific importins help disassemble the capsid
Herpesvirus • After fusion the tegument (most of it) is shed phosphorylation dependent • Contains NLSs on its capsids, binds microtubules via dynein • The virus is therefore transported as far as the pore. • It docks to the nuclear pore and then undergoes final disassembly, and the DNA is “injected” into the nucleus Note the capsid is “empty” - no dark center on EM From Whittaker Trends Microbiol 6: 178
Influenza virus • • • The nucleoprotein (NP) contains NLSs and the RNPs are small enough to translocate across the nuclear pore The key to influenza nuclear import is the p. H-dependent dissociation of the matrix protein (M 1) from the v. RNPs. This relies of the M 2 ion channel in the virus envelope, the target of amantadine From Whittaker Exp. Rev. Mol. Med. 8 February, http: //wwwermm. cbcu. cam. ac. uk/01002447 h. htm
Retroviruses • Simple + complex • • Simple retroviruses (oncoretroviruses) can only replicate in dividing cells, e. g. Rous sarcoma virus (RSV), avian leukosis virus (ALV). Nuclear entry occurs upon mitosis - the nuclear envelope breaks down and the virus is “passively” incorporated into the new nucleus This is relatively inefficient and restrictive for virus tropism Complex retroviruses (lentiviruses) have evolved mechanism for nuclear entry in non-dividing cells, e. g. HIV