Uvodna znanja Ispitna pitanja n n n n


znanja Matematika Fizička hemija Organska hemija Biohemija proteina i nukleinskih kiselina Struktura proteina")
 i sekvenca")
 n 4 b antiparalelne ravni (Greek key, meander)")
 stuktura n n 3 D struktura i 4 klase proteina; “pakovanje”")


 n n")


















? Introni i")











 E 1 Tiamin pirofosfat TPP Dihidrolipoil transacetilaza")


- Slides: 59
Uvodna znanja
Ispitna pitanja n n n n 1. Odredjivanje 3 D strukture proteina – kristalografija x-zracima 2. Odredjivanje 3 D strukture protina – difrakcija neutrona 3. Odredjivanje 3 D strukture proteina – NMR metode 4. Motivi ili supersekundarne strukture 5. Raznolikost proteina - Introni, egzoni, inteini i egzeini 6. Divergnetna evolucija familija proteina 7. Konvergentna evolucija 8. TIM barrel proteini 9. Evolucija proteina fuzijom genskih fragmenata 10. Enzimi sa više aktivnosti i nekovalentno udruživanje rezličitih aktivnosti 11. ”Arom” kompleks 12. Kompleks piruvat dehidrogenaze 13. Metode za odredjivanje stukture enzim-supstrat 14. Nomenklatura enzima
Uvodna (pred)znanja Matematika Fizička hemija Organska hemija Biohemija proteina i nukleinskih kiselina Struktura proteina
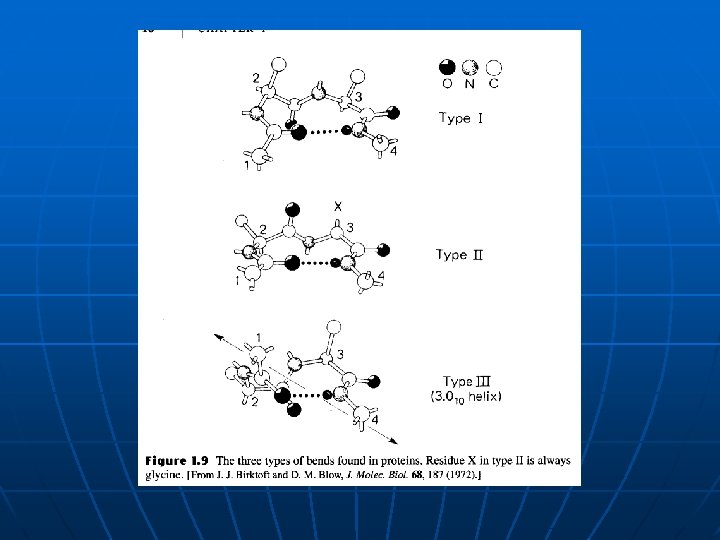
Struktura proteina n n Primarna struktura proteina – aminokiseline (21 proteinska aminokiselina) i sekvenca Posttranslacione modifikacije proteina - raznolikost Osnovni gradivni blokovi – peptidna veza (prolin, specifična aminokiselina) Sekundarna struktura (a heliks i b nabrana ravan). Amfipatična priroda heliksa, dipol ide paralelno sa osom heliksa (pozitivan pol je na N-kraju). n b-zavijutak (b-bend): 4 ostatka u nizu formiraju zavijutak kojim glavni polipeptidni lanac naglo menja pravac za 180 o. Na ovaj način je moguće povezati dve antiparalelne nabrane ravni. Tip I, II i III (čijim ponavljam nastaje 310 tip heliksa sa 3. 0 ostataka po okretu). n Ramachandran-ov dijagram
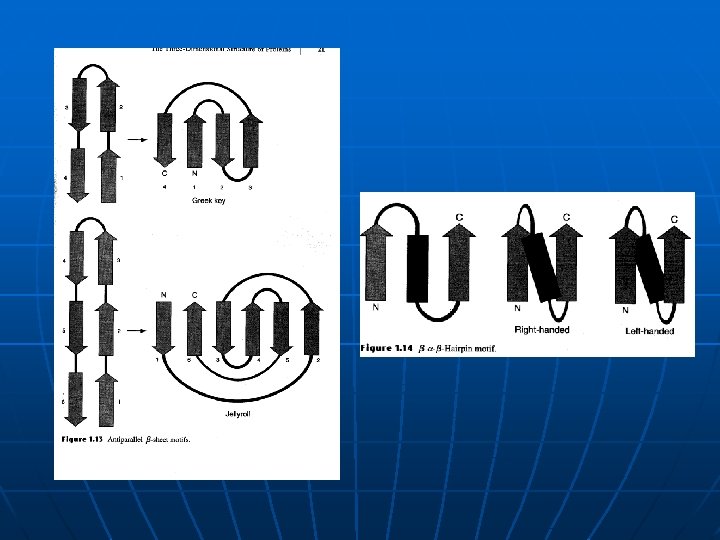
Motivi ili supersekundarne strukture b-ukosnica (b-hairpin) n 4 b antiparalelne ravni (Greek key, meander) n jellyroll (6 b-ravni) n Povezivanje paralelnih b-nabranih ravni nije tako jednostavno – konektor treba da promeni pravac dva puta. Strukture koje nastaju imaju sledeće topologije: b -a-b, b-b-b i češće su organizovanje po pravilu desne ruke. n Neuredjeni delovi (petlje) se često nalaze na površini i mogu da imaju vrlo važnu funkciju n Generalna pravila za povezivanje sekundarnih struktura u proteinima u motive tj. supersekundarne strukture: 1. Delovi sekundarnih struktura koji su bliski po sekvenci, često su u kontaktu i u 3 D 2. b-X-b struktura je češće organizovana po pravilu DR, nego po pravilu LR 3. Veze medju sek. str. ne prave čvorove, niti se ukrštaju. n Jedinica u strukturi globularnih proteina - domen i pravilo interakcije
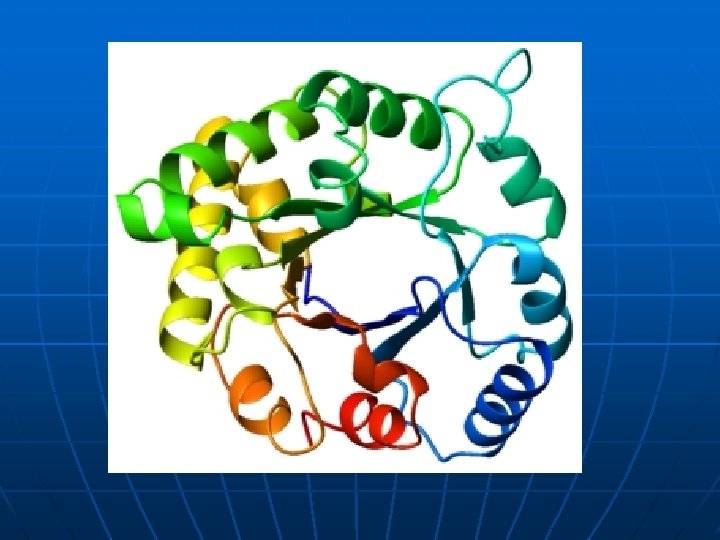
Trodimenzionalna (3 D) stuktura n n 3 D struktura i 4 klase proteina; “pakovanje” sekundarnih struktura. Kvaternerna struktura
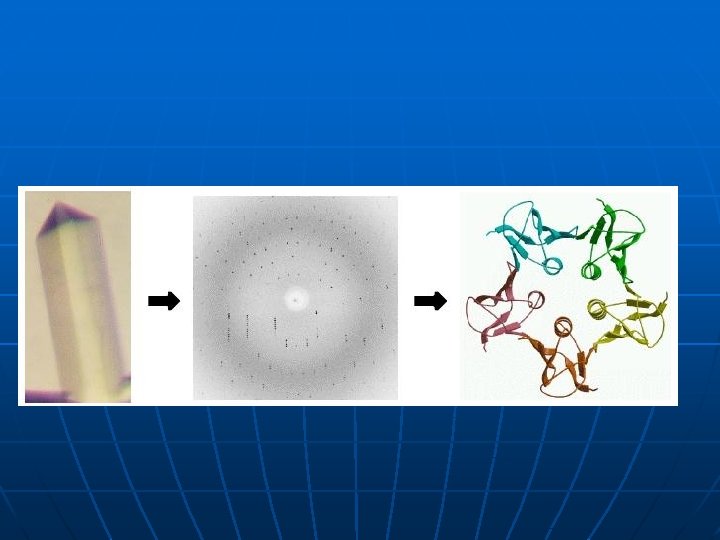
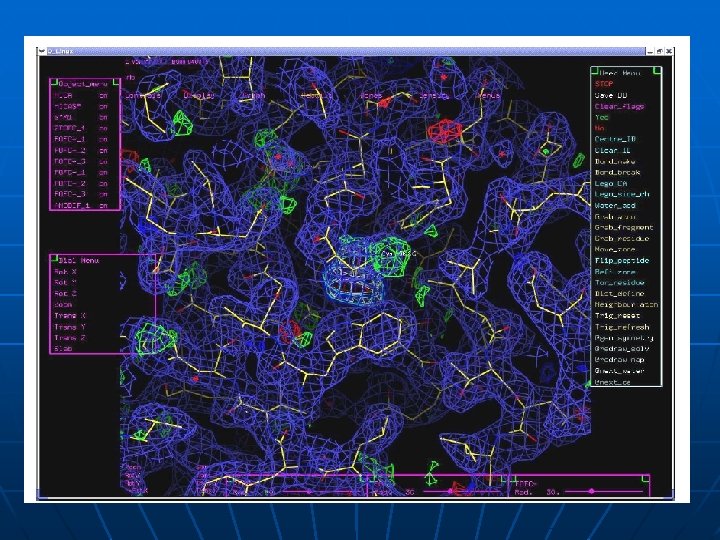
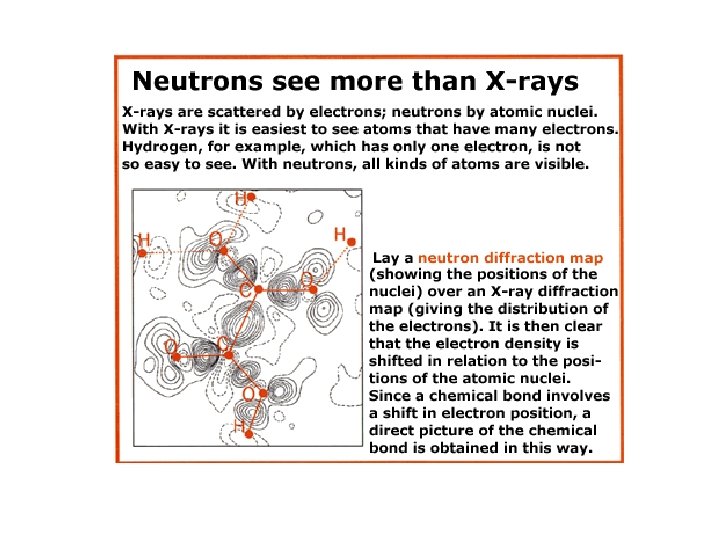
Metode za odredjivanje 3 D strukture proteina 1. x-ray kristalografija (difrakcija x-zraka na kristalisanim proteinima); isomorphous replacement, molecular replacement 2. Difrakcija neutrona (locira atome vodonika i važan je metod za rasvetljavanje reakcionih mehanizama) 3 - NMR u rastvoru 4. Small angle x-ray light scattering 5. Nisko rezolutne metode za odredjivanje topologije površine proteina masenom spektrometrijom 6. Predvidjanje 3 D strukture na osnovu sekvence proteina ili DNK
Vidljiva svetlost – talasna duzina: stotinak nm x-ray – 0. 1 nm
Amplifikacija signala n Fazni problem n Podesavanje Validacija (Ramachandranov dijagram) n n
Prakticni detalji
Kristalizacija
Metode za odredjivanje 3 D strukture proteina 1. x-ray kristalografija (difrakcija x-zraka na kristalisanim proteinima); isomorphous replacement, molecular replacement 2. Difrakcija neutrona (locira atome vodonika i važan je metod za rasvetljavanje reakcionih mehanizama) 3 - NMR u rastvoru 4. Small angle x-ray light scattering 5. Nisko rezolutne metode za odredjivanje topologije površine proteina masenom spektrometrijom 6. Predvidjanje 3 D strukture na osnovu sekvence proteina ili DNK
Difrakcija neutrona
Difrakcija neutrona
Metode za odredjivanje 3 D strukture proteina 1. x-ray kristalografija (difrakcija x-zraka na kristalisanim proteinima); isomorphous replacement, molecular replacement 2. Difrakcija neutrona (locira atome vodonika i važan je metod za rasvetljavanje reakcionih mehanizama) 3 - NMR u rastvoru 4. Small angle x-ray light scattering 5. Nisko rezolutne metode za odredjivanje topologije površine proteina masenom spektrometrijom 6. Predvidjanje 3 D strukture na osnovu sekvence proteina ili DNK
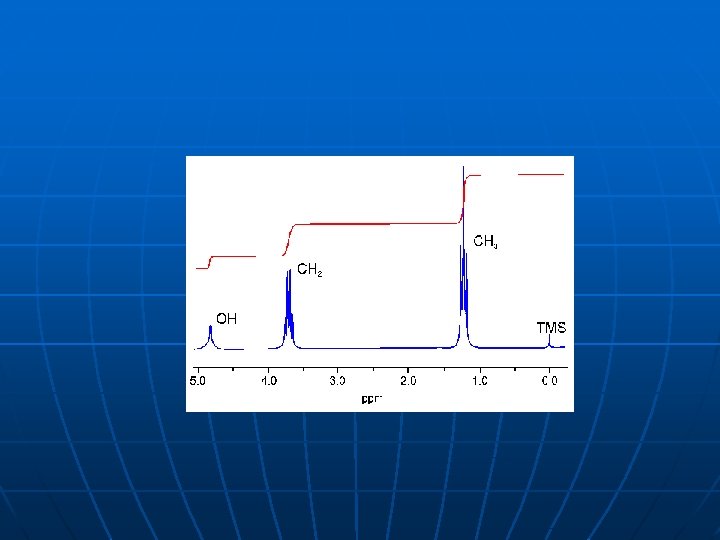
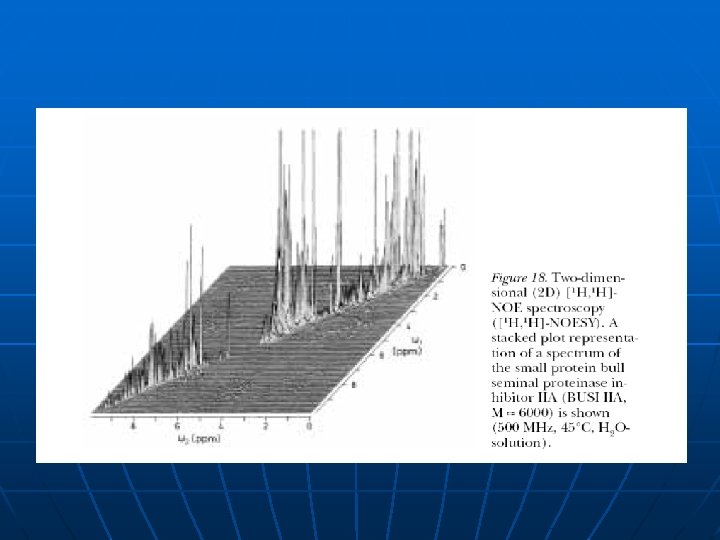
1 D NMR proteina
Metode za odredjivanje 3 D strukture proteina 1. x-ray kristalografija (difrakcija x-zraka na kristalisanim proteinima); isomorphous replacement, molecular replacement 2. Difrakcija neutrona (locira atome vodonika i važan je metod za rasvetljavanje reakcionih mehanizama) 3 - NMR u rastvoru 4. Small angle x-ray light scattering 5. Nisko rezolutne metode za odredjivanje topologije površine proteina masenom spektrometrijom 6. Predvidjanje 3 D strukture na osnovu sekvence proteina ili DNK
n Kvaternerna struktura – nekovalentno povezani oligomeri n Metode za odredjivanje kvaternerne strukture n n n Da li se na osnovu sekvence može predvideti katalitička funkcija enzima (više R ostataka ima katalitički potencijal) i značajne a. k. moraju da budu u odredjenom prostornom rasporedu tj. na odredjenoj udaljenosti da bi obavile katalitizu. Zašto su molekuli enzima relativno veliki? Da li su promene u sekvenci letalne za katalitičku funkciju?
Struktura kompleksa enzim-supstrat n n n Vezivanje malih molekula za protein u kristalu, molecular replacement za enzime čija 3 D struktura je odredjena Dobijanje stabilnih kompleksa Enzim-Supstrat: inhibitori, analozi supstrata, supstrati, ali u nereaktivnim uslovima Nove kristalografske tehnike (time-resolved crystalography). von Laue metod
Fleksibilnost i konformaciona pokretljivost proteina n n n Da li su kristalna struktura i struktura enzima u rastvoru identične? Rotacija u rastvoru “Disanje” regiona u proteinu Rotacija bočnih ostataka Kretanje domena – fleksibilnost segmenata
n http: //www. chem. ucsb. edu/~molvis ual/ABLE/induced_fit/index. html
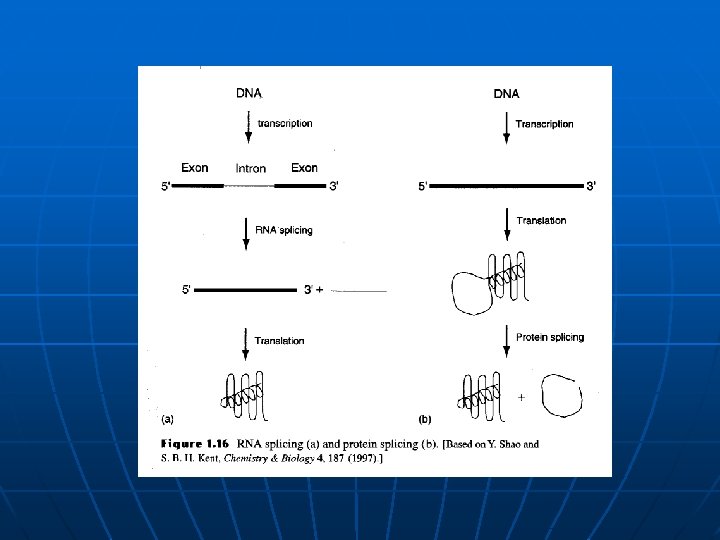
Raznolikost i evolucija proteina n n n Gen = Protein (produkt gena)? Introni i egzoni (ribozimi); obrada RNA, alternativna obrada RNA Inteini i eksteini; protein splicing
Divergentna i konvergentna evolucija proteinskih familija Serin proteaze su primer divergentne evolucije i verovatno su nastale od zajedničkog pretka. Homologi proteini Konvergentna evolucija proteina – različita 3 D struktura, ali isti mehanizam delovanja Endopeptidaza termolizin karboksipeptidaze i
Konvergentno ili divergentno? n 1. 2. 3. 4. 5. 6. Kriterijumi za testiranje – da li su dva proteina evoluirala od zajedničkog pretka? Sličnost DNK sekvenci Sličnost amino kiselinskih sekvenci Sličnost 3 D struktura Interakcije Enzim-Supstrat su slične Katalitički mehanizmi su slični Sekvenca fragmenata polipeptidnog niza koji su esencijalni za katalizu je očuvana.
TIM barrel proteini
Evolucija proteina fuzijom genskih fragmenata n n n n n Domenska struktura NAD+-zavisnih dehidrogenaza: gen koji vezuje nukleotid + gen koji kodira katalitički domen Domen kompaktna strukturna jedinica Da li su egzoni funkcionalne jedinice u evoluciji proteina? Evolucija fuzijom fragmenata se odvijala preko osnovnih jedinica “domena”. Razmeštanje domena in vitro (E. coli aspartat transkarbamoilaza (ATCaza)) ima dva globularna domena. E. coli ornitin transkarbamoilaza ima 32 % identičnu sekvencu. Oligomerizacija razmenom domena (domen swapping). Intra i intemolekularno uparivanje domena (oligomerni proteini). Homologija, sličnost i identičnost sekvence
Viši nivoi organizacije enzimskih aktivnosti
Viši nivoi organizacije: multienzimski kompleksi n n n Enzimi uključeni u metaboličke procese koji su u sekvenci , ili složene biohemijske procese su često organizovani u fizičke agregate: triptofan sintetaza, sintetaza masnih kiselina, primozom ribozomi.
Enzimi sa više aktivnih centara i nekovalenta asocija različitih aktivnosti - Triptofan sintaza, a 2 b 2 tetramer i nalazi se u okviru triptofanskog operona Indol-3 -glicerol fosfat + serin ------> Trp + gliceraldehid-3 fosfat (a 2 b 2) Indol-3 -glicerol fosfat ------> indol + gliceraldehid-3 -fosfat (a) indol + serin ------> Trp + gliceraldehid-3 -fosfat (a 2 b 2) Podjedinice u komplesku su povezane 25 -30 Å dugačkim tunelom.
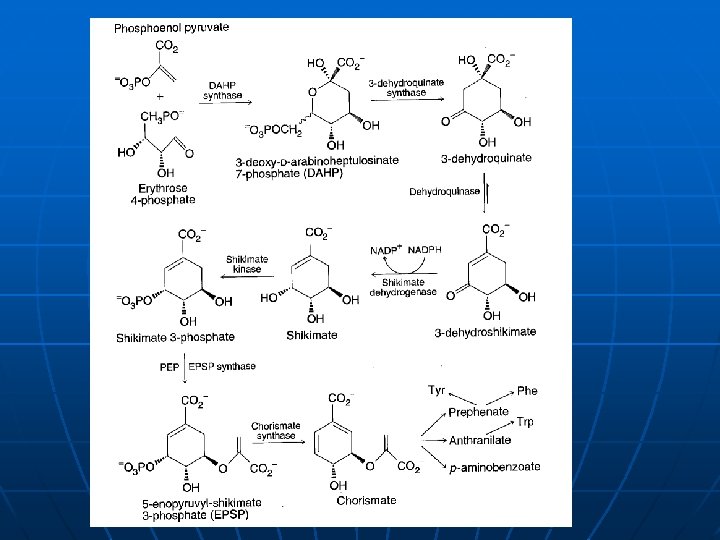
Enzimi sa više aktivnih centara - - - Indol-3 -glicerol fosfat-sintazafosforibozil antranilat izomeraza (bifunkcionalni enzim). Genetskim inženjeringom razdvojene podjedinice imaju istu katalitičku aktivnost. ? Arom kompleks (šikamat mehanizam biosinteze aromatičnih jedinjenja), kompleks piruvat dehidrogenaze
Enzimi sa više aktivnih centara E. coli n Arom kompleks (šikamat mehanizam biosinteze aromatičnih jedinjenja) Saccharomyces cerevisiae, N. crassa 5 5 aktivnih centara u monofunkcionalnih jednom polipeptidnom enzima nizu
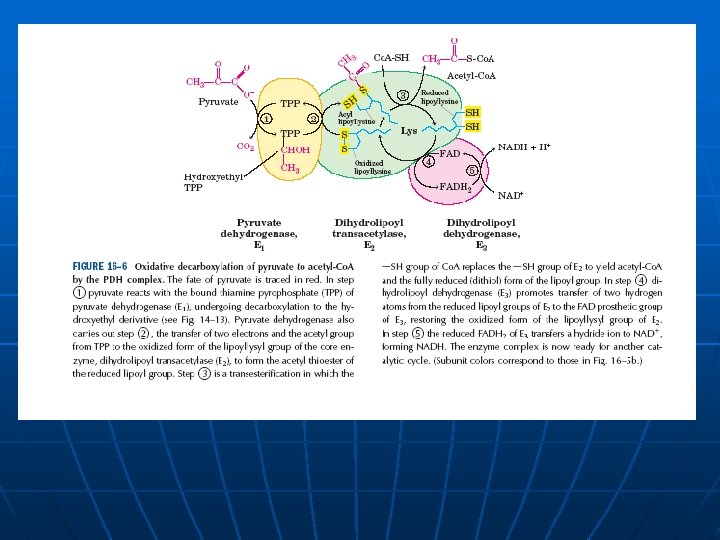
Zašto multienzimski kompleksi? n n n Poboljšana kataliza: smanjuje se vreme difuzije intermedijera u povezanim procesima od jednog do drugog enzima Kanalisanje supstrata Zaštita hemijski reaktivnih intermedijera od izlaganja vodenoj sredini Servis (u slučaju piruvat dehidrogenaze, jedna podjedinica može da prosledi reagens (servisira) ostale podjedinice) Kooridinisana regulacija Koordinisana ekspresija
Kompleks piruvat dehidrogenaze
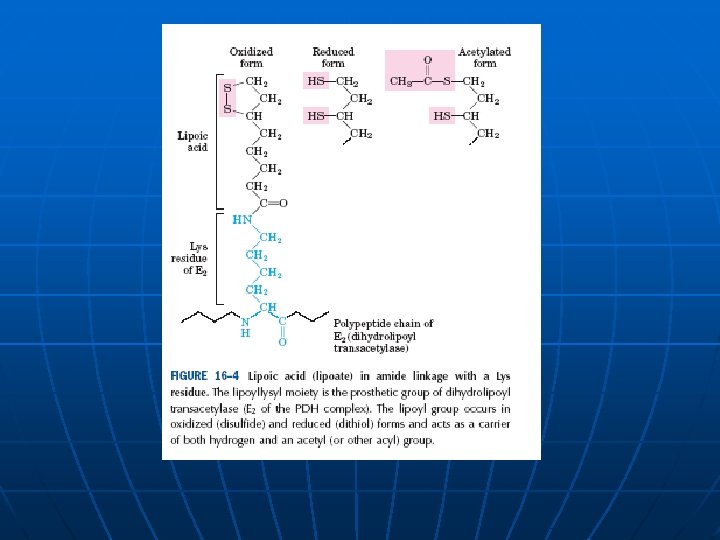
Enzim Prosteticna grupa Piruvat dehidrogenaza (a 2) E 1 Tiamin pirofosfat TPP Dihidrolipoil transacetilaza E 2 Lipoamid Dihidrolipoil dehidrogenaza (a 2) E 3 FAD
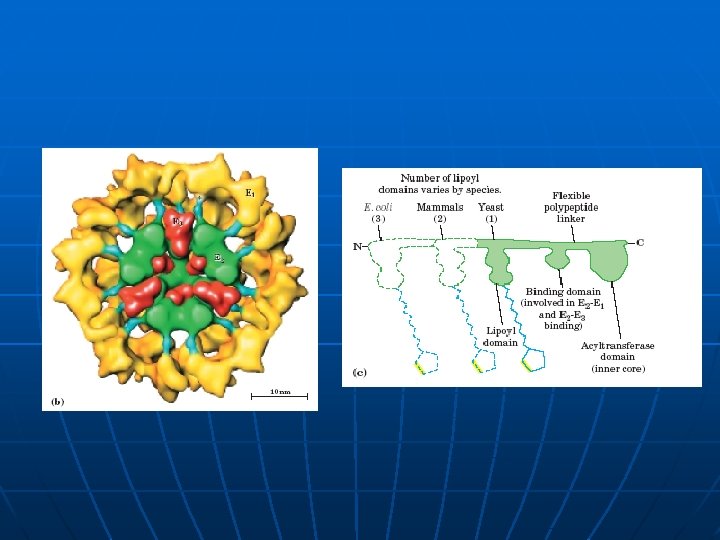
n Sectioned schematic view of an icosahedral pyruvate dehydrogenase complex based on electron cryomicroscopic analysis of an E 1 E 2 sub-complex from B. stearothermophilus. Three of the 60 E 2 molecules (colored red, green and yellow) are highlighted. The movement of the swinging E 2 lipoyl domain in the annular region between the inner core (cyan) of E 2 molecules and the outer shell of E 1 molecules (purple) is proposed to be a critical feature underlying active site coupling in the complex.
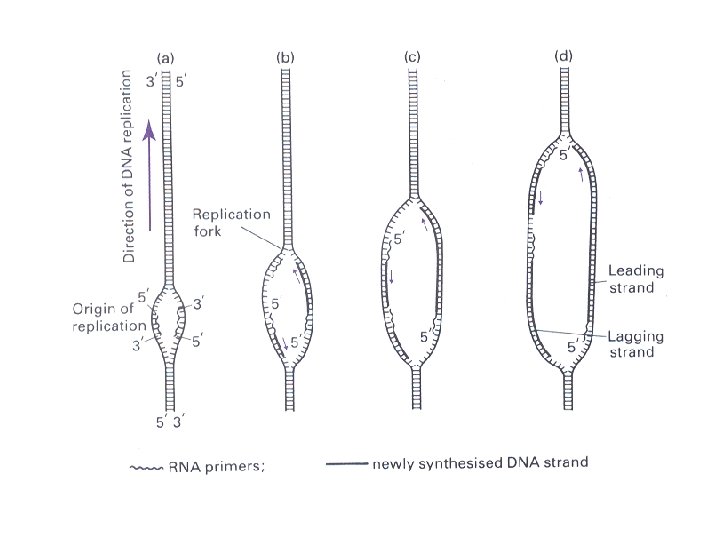
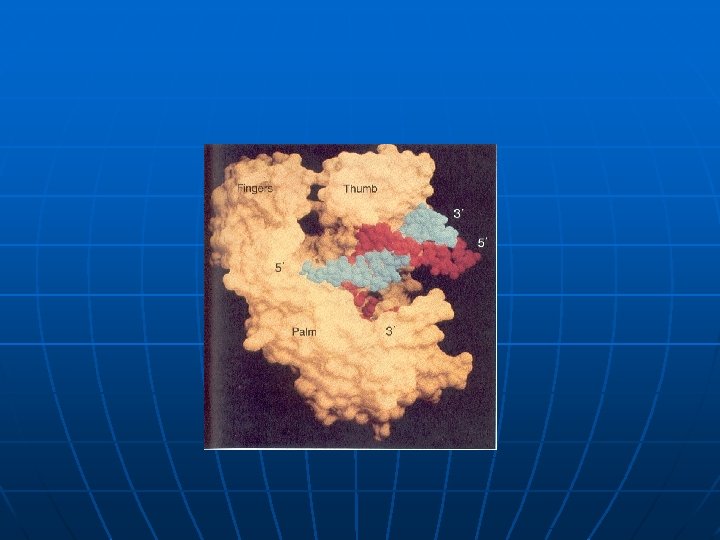
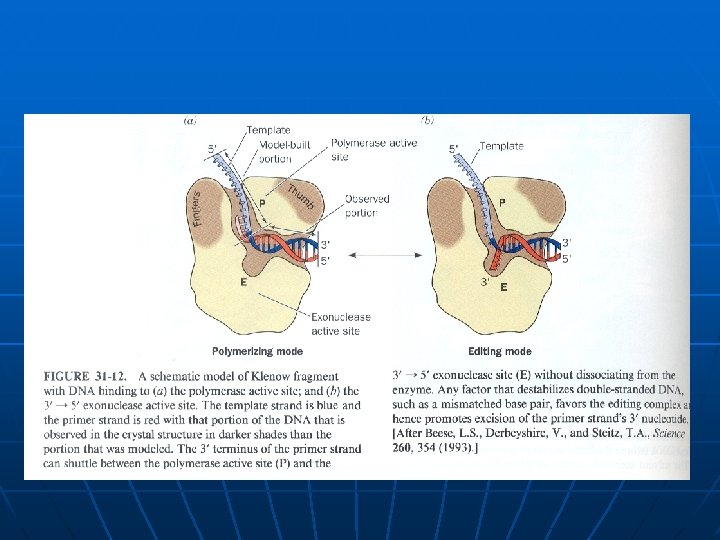
Multifunkcionalni enzimi – DNK polimeraze n n n DNK polimeraza I E. coli (928 a. k. ) 324 -518 polimerizacija lanca DNK 1 -320 5’->3’ egzonukleazna aktivnost 520 -928 3’->5’ egzonukleazna aktivnost DNK polimeraza (reverzna transkriptaza) HIV virusa, ima i ribonukleazni domen kojim iz DNK-RNK hibrida uklanja RNK tokom procesa replikacije RNK genoma virusa.