Using molecular replacement to exploit multiple crystal forms










")

 Difficult structure from Bogos Agianian (Piet")

")
")





















- Slides: 38
Using molecular replacement to exploit multiple crystal forms Randy Read, Airlie Mc. Coy & Gábor Bunkóczi Tom Terwilliger
Solving the structure of angiotensinogen • Human: 1 crystal form • • Rat: 2 crystal forms • • • 3. 3Å, 1 copy, P 41212 2. 8Å, 2 copies, C 2 3. 15Å, 2 copies, P 3221 Mouse: 2 crystal forms • • 2. 1Å, 1 copy, P 6122 2. 95Å, 4 copies, C 2221
Human angiotensinogen: molecular replacement human
Human angiotensinogen: molecular replacement human
Human angiotensinogen: molecular replacement human
Human angiotensinogen after MR MR model Final model map CC = 0. 38
Human angiotensinogen after MR+DM MR model Final model map CC = 0. 50
Solving angiotensinogen structures human rat C 2 rat P 3221 + Gd. Cl 3
Rat C 2 angiotensinogen after MR with density + 2 -fold averaging MR model Final model map CC = 0. 44 (0. 48 with Gd. Cl 3)
Rat C 2 angiotensinogen after 4 -fold multicrystal averaging MR model Final model map CC = 0. 53 (0. 59 with Gd. Cl 3)
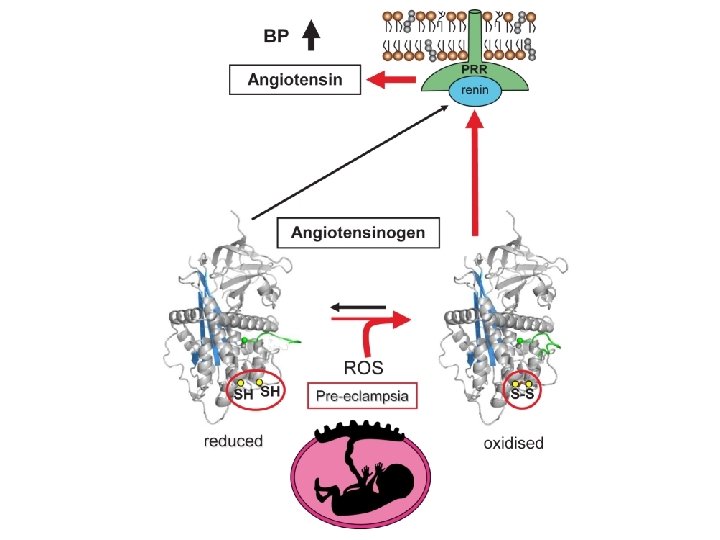
Renin: angiotensinogen complex Renin S-S (18 -138)
Solving Drosophila GST 2 (1 M 0 U) Difficult structure from Bogos Agianian (Piet Gros) • Find one of two copies with ensemble of 3 structures (28 -30% identity) • • • search for second copy fails Find second copy as density from first • • this succeeds: TFZ=10. 0 trick to doing this: assign low error to first copy, higher error to second
How does averaging add information? • Nyquist-Shannon sampling theorem: • doubly-sampled Fourier transform can be reconstructed perfectly
Molecule and molecular transform (Images courtesy of Kevin Cowtan)
Crystal diffraction samples molecular transform on reciprocal lattice (Images courtesy of Kevin Cowtan)
Non-isomorphism changes sampling of molecular transform Cell change Rotated molecule (Images courtesy of Kevin Cowtan)
Absolute vs. relative cell change d=6Å d=3Å b=27Å b=24Å a=48Å a=51Å
“Protein X” 525 residues • Only distant homologues in PDB • • eight models with sequence identity about 20%
Ensemble of 8 possible models
Molecular replacement trials • MR with individual models failed • • complete or trimmed with FFAS server MR with ensemble failed initially • but now works with new “trim” option in phenix. ensembler
Trimming untrimmed
Trimming trimmed
Data sets • Space group P 3112 Crystal Native Gd. Cl 3 Na. I a =b 70. 9Å 68. 8Å 72. 5Å c 286. 0Å 289. 0Å 286. 4Å dmin 2. 4Å 3. 3Å 3. 2Å(3. 6Å) ΔBaniso λ 35Å2 0. 9728Å 96Å2 1. 475Å 92Å2 0. 9763Å
Experimental phasing with Gd. Cl 3 • Gd. Cl 3 derivative diffracts to about 3. 3Å • • • good anomalous signal (phenix. xtriage) to 4. 3Å solve substructure with phenix. hyss (4 sites) phase with Phaser solvent flattening see some features of fold, not complete trace
Real-space MR against Gd. Cl 3 map • Use real-space MR to place models in density • cut out density from SAD map • • back-transform, treat as observed data rotation search, phased translation Resolution, phases too poor to rebuild • Model provides starting mask •
Real-space MR model in Gd density
Solve native crystal with Gd. Cl 3 density Use real-space MR model to construct envelope • Cut out Gd. Cl 3 density for one molecule • • place in large unit cell (4 x extent in each direction) FFT to get molecular transform Use density for MR solution of native crystal • • RFZ=3. 8, TFZ=22. 9 or just rigid-body refinement starting from identity operator
Solve and phase iodide soak • Diffracts to 3. 2Å, but data sparse beyond 3. 6Å • • good anomalous signal only to 5. 4Å (phenix. xtriage) Ab initio substructure determination failed • phenix. hyss Rigid-body refinement of Gd. Cl 3 density model • Use Fc from MR solution as partial model for MR -SAD phasing • • SAD log-likelihood-gradient maps yield 11 sites
Multi-crystal averaging Get operators from transformations applied to Gd. Cl 3 density • Starting maps from SAD phasing (derivatives) or MR with Gd. Cl 3 density (native) • Carried out with phenix. multi_crystal_average •
Morphing • Use phenix. morph_model to morph real-space MR model into native averaged density • correlation increases from 0. 296 to 0. 338
Initial model-building • Build with phenix. autobuild into averaged native map, start from morphed MR model • • 364 residues, 177 assigned to sequence R=0. 42, Rfree=0. 48
Iterative averaging and rebuilding • Carry on with iterative improvement • • • MR-SAD phasing with current model multi-crystal averaging Auto. Build, do some manual rebuilding MR-SAD phasing with updated model multi-crystal averaging Iteration has improved anomalous substructures • • Gd. Cl 3: 4 sites to 8 sites (2 split) Na. I: 11 sites to 14 sites
Map after iterative process MR model Current model
Tools • Cutting out density • • • Molecular replacement with density • • • Molrep Phaser SAD phasing starting from density • • phenix. cut_out_density (Tom Terwilliger) cmapcut (Kevin Cowtan) Phaser SAD LLG, giving density as partial model Multi-crystal averaging • • dmmulti phenix. multi_crystal_average
Acknowledgements • Phaser: • • Angiotensinogen: • • • Airlie Mc. Coy, Gabor Bunkoczi Penny Stein, Robin Carrell, Aiwu Zhou Mike Murphy, Fiona Broughton Pipkin “Protein X” • Mykhaylo Demydchuk, Aiwu Zhou, Janet Deane, Penny Stein
The PHENIX Project Lawrence Berkeley Laboratory Paul Adams, Ralf Grosse. Kunstleve, Pavel Afonine, Nat Echols, Nigel Moriarty, Jeff Headd, Nicholas Sauter, Peter Zwart Randy Read, Airlie Mc. Coy, Gabor Bunkoczi, Rob Oeffner Cambridge University An NIH/NIGMS funded Program Project Los Alamos National Laboratory Tom Terwilliger, Li-Wei Hung Duke University Jane & David Richardson, Vincent Chen, Chris Williams, Bryan Arendall, Swati Jain, Bradley Hintze