TRANSPORT OVERVIEW 3 LEVELS OF TRANSPORT OCCUR IN

 UPTAKE AND RELEASE OF")




-units of water potential pressure • PURE WATER IN AN")





















 IN XYLEM,")















- Slides: 45
TRANSPORT OVERVIEW 3 LEVELS OF TRANSPORT OCCUR IN PLANTS: 1) UPTAKE AND RELEASE OF WATER AND SOLUTES BY INDIVIDUAL CELLS 2) SHORT-DISTANCE CELL-TO-CELL TRANSPORT AT THE LEVEL OF TISSUES AND ORGANS 3) LONG-DISTANCE TRANSPORT OF SAP IN XYLEM AND PHLOEM AT THE WHOLEPLANT LEVEL
TRANSPORT AND MEMBRANES • THE PLASMA MEMBRANE’S SELECTIVE PERMEABILITY CONTROLS THE MOVEMENT OF SOLUTES BETWEEN A PLANT CELL AND THE EXTRACELLULAR FLUIDS. • TRANSPORT PROTEINS MAY FACILITATE DIFFUSION BY SERVING AS CARRIER PROTEINS OR FORMING SELECTIVE CHANNELS
PROTON PUMPS • A PROTON PUMP HYDROLYZES ATP AND USES THE ENERGY TO PUMP HYDROGEN IONS (H+) OUT OF THE CELL – THIS PRODUCES A PROTON GRADIENT WITH A HIGHER CONCENTRATION OUTSIDE OF THE CELL – PRODUCES A MEMBRANE POTENTIAL, SINCE THE INSIDE OF THE PLANT CELL IS NEGATIVE IN RELATION TO THE OUTSIDE • THIS MEMBRANE POTENTIAL AND THE STORED ENERGY OF THE PROTON GRADIENT ARE USED BY THE PLANT TO TRANSPORT MANY DIFFERENT MOLECULES • POTASSIUM (K+) IONS ARE PULLED INTO THE CELLS BECAUSE OF THE ELECTROCHEMICAL GRADIENT • **THE INVOLVEMENT OF PROTON PUMPS IN THE TRANSPORT PROCESSES OF PLANT CELLS IS A SPECIFIC APPLICATION OF CHEMIOSMOSIS
SOLUTE TRANSPORT IN PLANT CELLS
WATER POTENTIAL DRIVES TRANSPORT • OSMOSIS RESULTS IN THE NET UPTAKE OR LOSS OF WATER BY THE CELL AND DEPENDS ON WHICH COMPONENT, THE CELL OR EXTRACELLULAR FLUIDS, HAS THE HIGHEST WATER POTENTIAL • WATER POTENTIAL (Y) = THE FREE ENERGY OF WATER THAT IS A CONSEQUENCE OF SOLUTE CONCENTRATION AND APPLIED PRESSURE; PHYSICAL PROPERTY PREDICTING THE DIRECTION WATER WILL FLOW
WATER POTENTIAL • MEGAPASCALS (Mpa)-units of water potential pressure • PURE WATER IN AN OPEN CONTAINER HAS A Y OF ZERO Mpa (Y=O) • ADDITION OF SOLUTES TO WATER LOWERS THE Y INTO THE NEGATIVE RANGE • INCREASED PRESSURE RAISES THE Y INTO THE NEGATIVE RANGE
QUANTITATIVE WATER POTENTIAL • THE EFFECTS OF PRESSURE AND SOLUTE CONCENTRATION ON WATER POTENTIAL ARE REPRESENTED BY Y =YP + YS YP = PRESSURE POTENTIAL YS = SOLUTE POTENTIAL OR OSMOTIC POTENTIAL
WATER POTENTIAL AND WATER MOVEMENT
WATER RELATIONS OF PLANT CELLS
AQUAPORINS • AQUAPORINS ARE WATER-SPECIFIC CHANNELS MADE UP OF TRANSPORT PROTEINS, HAVE BEEN DISCOVERED IN PLANT AND ANIMAL CELLS • AQUAPORINS DO NOT ACTIVELY TRANSPORT WATER, BUT RATHER FACILITATE ITS DIFFUSION
TONOPLAST • TONOPLAST = MEMBRANE SURROUNDING THE LARGE CENTRAL VACUOLE FOUND IN PLANT CELLS; IMPORTANT IN REGULATING INTRACELLULAR CONDITIONS • CONTAINS INTEGRAL TRANSPORT PROTEINS THAT CONTROL THE MOVEMENT OF SOLUTES BETWEEN THE CYTOSOL AND THE VACUOLE • HAS A MEMBRANE POTENTIAL; PROTON PUMPS IN THE TONOPLAST HELP THE PLASMA MEMBRANE MAINTAIN A LOW H+ CONCENTRATION IN THE CYTOSOL BY MOVING H+ INTO THE VACUOLE
PLASMODESMATA • PLASMODESMATA CONNECT THE CYTOSOLIC COMPARTMENTS OF NEIGHBORING CELLS; THIS CYTOPLASMIC CONTINUUM IS CALLED THE SYMPLAST • THE WALLS OF ADJACENT CELLS ARE CONNECTED TO FORM A CONTINUUM OF CELL WALLS CALLED APOPLAST
COMPARTMENTS OF PLANT CELLS THE SYMPLAST & APOPLAST BOTH LATERALLY TRANSPOR
BULK FLOW • THIS TYPE OF TRANSPORT IS USUALLY ALONG THE VERTICAL AXIS OF THE PLANT (UP & DOWN) FROM THE ROOTS TO THE LEAVES AND VICE VERSA • VASCULAR TISSUES ARE INVOLVED IN THIS TYPE OF TRANSPORT AS DIFFUSION WOULD BE TOO SLOW • BULK FLOW (MOVEMENT DUE TO PRESSURE DIFFERENCES) MOVES WATER AND SOLUTES THROUGH XYLEM VESSELS AND SIEVE TUBES • TRANSPIRATION REDUCES PRESSURE IN THE LEAF XYLEM; THIS CREATES A TENSION WHICH PULLS SAP UP THROUGH THE XYLEM FROM THE ROOTS
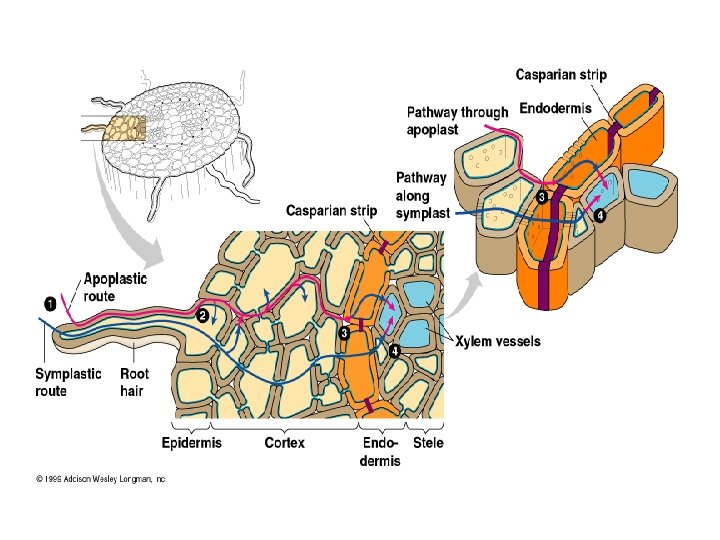
ABSORPTION BY ROOTS • WATER AND MINERALS ENTER PLANTS THROUGH THE FOLLOWING PATHWAY SOIL>>>>EPIDERMIS>>>ROOT CORTEX>>>XYLEM
LATERAL TRANSPORT OF MINERALS AND WATER IN ROOTS
VIDEO: TRANSPORT
WATER AND MINERAL ABSORPTION SOIL EPIDERMIS -MOST ABSORPTION OCCURS NEAR ROOT TIPS WHERE THE EPIDERMIS IS PERMEABLE TO WATER -ROOT HAIRS, EXTENSIONS OF EPIDERMAL CELLS, INCREASE THE SURFACE AREA AVAILABLE FOR ABSORPTION -MOST PLANTS FORM SYMBIOTIC RELATIONSHIPS WITH FUNGI; THE “INFECTED” ROOTS FORM MYCORRHIZAE, A STRUCTURE MADE FROM THE PLANT ROOTS AND THE HYPHAE OF FUNGI. WATER AND MINERALS ABSORBED BY THE HYPHAE ARE TRANSFERRED TO THE HOST PLANT
EPIDERMIS ROOT CORTEX: -LATERAL TRANSPORT OF MINERALS AND WATER THROUGH THE ROOT IS USUALLY BY A COMBINATION OF APOPLASTIC AND SYMPLASTIC ROUTES -THE APOPLASTIC ROUTE EXPOSES PARENCHYMAL CORTEX CELLS TO SOIL SOLUTION -THE SYMPLASTIC ROUTE MAKE SELECTIVE MINERAL ABSORPTION POSSIBLE
ENDODERMIS FUNCTIONS ROOT CORTEX XYLEM: • ONLY MINERALS USING THE SYMPLASTIC ROUTE MAY MOVE DIRECTLY INTO THE VASCULAR TISSUES. • MINERALS AND WATER PASSING THROUGH APOPOLASTS ARE BLOCKED AT THE ENDODERMIS BY A CASPERIAN STRIP (A RING OF SUBERIN AROUND EACH CELL IN THE ENDODERMIS) AND MUST ENTER AN ENDODERMAL CELL • WATER AND MINERALS ENTER INTO THE STELE THROUGH THE CELLS OF THE ENDODERMIS
TRANSPORT OF XYLEM SAP • THE ASCENT OF XYLEM SAP DEPENDS MAINLY ON TRANSPIRATION AND THE PHYSICAL PROPERTIES OF WATER • THE SHOOT DEPENDS UPON AN EFFICIENT DELIVERY OF ITS WATER SUPPLY – XYLEM SAP FLOWS UPWARD AT 15 m PER HOUR OR FASTER – XYLEM VESSELS ARE CLOSE TO EACH LEAF CELL, BECAUSE VEINS BRANCH THROUGHOUT THE LEAVES WATER TRANSPORTED UP FROM ROOTS MUST REPLACE THAT LOST BY TRANSPIRATION -TRANSPIRATION IS THE EVAPORATION OF WATER FROM THE AERIAL PARTS OF A PLANT -THE UPWARD FLOW OF XYLEM SAP ALSO PROVIDES NUTRIENTS (MINERALS) TO THE SHOOT SYSTEM
ROOT PRESSURE • WHEN TRANSPIRATION IS LOW, ACTIVE TRANSPORT OF IONS INTO THE XYLEM DECREASES THE STELE’S WATER POTENTIAL AND CAUSES WATER FLOW INTO THE STELE. THIS OSMOTIC WATER UPTAKE INCREASES PRESSURE WHICH FORCES FLUID UP THE XYLEM (= ROOT PRESSURE) – ROOT PRESSURE CAUSES GUTTATION (EXUDATION OF WATER DROPLETS AT LEAF MARGINS) • ROOT PRESSURE IS NOT THE MAJOR MECHANISM DRIVING THE ASCENT OF XYLEM SAP – CANNOT KEEP PACE WITH TRANSPIRATION – CAN ONLY FORCE WATER UP A FEW METERS
PULLING XYLEM SAP TRANSPIRATION PULLS XYLEM SAP UPWARD, AND COHESION OF WATER TRANSMITS THE UPWARD-PULL ALONG THE ENTIRE LENGTH OF XYLEM
TRANSPIRATIONAL PULL
TRANSPIRATIONAL PULL • TRANSPIRATIONAL PULL DEPENDS UPON THE CREATION OF NEGATIVE PRESSURE • GASEOUS WATER IN DAMP INTERCELLULAR LEAF SPACES DIFFUSES INTO THE DRIER ATMOSPHERE THROUGH STOMATA • THE LOST WATER VAPOR IS REPLACED BY EVAPORATION FROM MESOPHYLL CELLS BORDERING THE AIRSPACES • THE REMAINING WATER FILM, ADHERING TO THE HYDROPHILIC CELL WALLS, RETREATS INTO THE CELL WALL PORES • COHESION IN THIS SURFACE FILM OF WATER RESISTS AN INCREASE IN THE SURFACE AREA OF THE FILM-A SURFACE TENSION EFFECT • THE WATER FILM FORMS A MENISCUS DUE TO THE NEGATIVE PRESSURE CAUSED BY THE ADHESION AND COHESION • THIS NEGATIVE PRESSURE PULLS WATER FROM THE XYLEM, THROUGH THE MESOPHYLL, TOWARD THE SURFACE FILM ON CELLS BORDERING THE STOMATA
MEASURING WATER POTENTIAL OF LEAVES
COHESION AND ADHESION OF WATER • THE TRANSPIRATIONAL PULL ON THE XYLEM SAP IS TRANSMITTED TO THE SOIL SOLUTION. COHESION OF WATER DUE TO H+ BONDS ALLOWS FOR THE PULLING OF WATER FROM THE TOP OF THE PLANT WITHOUT BREAKING THE “CHAIN” • THE ADHESION OF WATER (BY H+ BONDS) TO THE HYDROPHILIC WALLS OF XYLEM CELLS ALSO HELPS PULL AGAINST GRAVITY • THE SMALL DIAMETER OF VESSELS AND TRACHEIDS IS IMPORTANT TO THE ADHESION EFFECT
COHESION CON’T • THE UPWARD PULL OF SAP CAUSES TENSION (NEGATIVE PRESSURE) IN XYLEM, WHICH DECREASES WATER POTENTIAL AND ALLOWS PASSIVE FLOW OF WATER FROM SOIL INTO STELE • TRANSPIRATIONAL PULL CAN EXTEND DOWN TO THE ROOTS ONLY THROUGH AN UNBROKEN CHAIN OF WATER MOLECULES • CAVITATION (FOMATION OF A WATER VAPOR POCKET IN XYLEM) BREAKS THE CHAIN OF WATER MOLECULES AND THE PULL IS STOPPED
ASCENT OF XYLEM SAP
CONTROL OF TRANSPIRATION • TRANSPIRATION RESULTS IN A TREMENDOUS WATER LOSS FROM THE PLANT. THIS WATER IS REPLACED BY THE UPWARD MOVEMENT OF WATER THROUGH THE EXLEM. GUARD CELLS SURROUNDING STOMATA BALANCE THE REQUIREMENTS FOR PHOTOSYNTHESIS WITH THE NEED TO CONSERVE WATER.
TRANSPIRATION VIDEO
GUARD CELLS • GUARD CELLS = CELLS THAT FLANK STOMATA AND CONTROL STOMATAL DIAMETER BY CHANGING SHAPE • WHEN TURGID, GUARD CELLS “BUCKLE DUE TO RADIALLY-ARRANGED MICROFIBRILS AND STOMATA OPEN • WHEN FLACCID, GUARD CELLS SAG AND STOMATAL OPENINGS CLOSE
• THE CHANGE IN TURGOR PRESSURE THAT REGULATES STOMATAL OPENING RESULTS FROM REVERSIBLE UPTAKE AND LOSS OF K+ BY GUARD CELLS – UPTAKE OF K+ DECREASES GUARD CELL WATER POTENTIAL SO WATER IS TAKEN UP, CELLS BECOME TURGID, AND STOMATA OPEN. THE TONOPLAST PLAYS A ROLE AS MOST OF THE K+ AND WATER ARE STORED IN THE VACUOLE – THE INCREASE IN POSITIVE CHARGE IS COUNTERED BY THE UPTAKE OF CHLORIDE (Cl-), EXPORT OF H+ IONS RELEASED FROM ORGANIC ACIDS, AND THE NEGATIVE CHARGES ACQUIRED BY ORGANIC ACIDS AS THEY LOSE THEIR PROTONS – CLOSING OF THE STOMATA RESUTLS WHEN K+ EXITS THE GUARD CELLS AND CREATES AN OSMOTIC LOSS OF WATER
STOMATAL CONTROL
VIDEO: STOMATA AND GUARD CELLS
XEROPHYTES: ARID REGIONS • XEROPHYTES, PLANTS ADAPTED TO ARID CLIMATES, HAVE SOME OF THE FOLLOWING EVOLUTIONARY ADAPTATIONS THAT REDUCE TRANSPIRATION: – SMALL, THICK LEAVES (REDUCED SURFACE AREA: VOLUME, SO LESS WATER LOSS) – A THICK CUTICLE – STOMATA ARE DEPRESSIONS ON THE UNDERSIDE OF LEAVES TO PROTECT FROM WATER LOSS DUE TO DURING WINDS (LASERDISK) – SOME SHED LEAVES IN THE DRIEST TIME OF THE YEAR – CACTI AND OTHERS STORE WATER IN STEMS DURING THE WET SEASON
TRANSLOCATION OF PHLOEM SAP • TRANSLOCATION = THE TRANSPORT OF THE PRODUCTS OF PHOTOSYNTHESIS BY PHLOEM TO THE REST OF THE PLANT • PHLOEM TRANSLOCATES ITS SAP FROM SUGAR SOURCES TO SUGAR SINKS • SUGAR SOURCE = ORGAN WHERE SUGAR IS PRODUCES BY PHOTOSYNTHESIS OR BY THE BREAKDOWN OF STARCH • SUGAR SINK = ORGAN THAT CONSUMES OR STORES SUGAR
PHLOEM LOADING AND UNLOADING • SUGAR PRODUCES AT A SOURCE MUST BE LOADED INTO SIEVE-TUBE MEMBERS BEFORE IT CAN BE TRANSLOCATED TO A SINK • IN SOME PLANT SPECIES, THE SUGAR MAY MOVE THROUGH THE SYMPLAST FROM MESOPHYLL CELLS TO THE SIEVE MEMBERS • IN OTHER SPECIES, THE SUGAR USES A COMBINATION OF SYMPLASTIC AND APOPLASTIC ROUTES • SOME PLANTS HAVE TRANSFER CELLS. THESE ARE MODIFIED COMPANION CELLS WHICH HAVE NUMEROUS INGROWTHS OF THEIR WALLS
VIDEO: TRANSLOCATION
• IN PLANTS SUCH AS CORN, ACTIVE TRANSPORT ACCUMULATES SUCROSE IN SIEVE-TUBE MEMBERS TO TWO TO THREE TIMES THE CONCENTRATION IN MESOPHYLL CELLS – PROTON PUMPS POWER THIS TRANSPORT BY CREATING A H+ GRADIENT – A MEMBRANE PROTEIN USES THE POTENTIAL ENERGY STORED IN THE GRADIENT TO DRIVE THE COTRANSPORT OF SUCROSE BY COUPLING SUGAR TRANSPORT TO THE DIFFUSION OF H+ BACK INTO THE CELL
LOADING OF SUCROSE INTO PHLOEM
PRESSURE FLOW • PHLOEM SAP FLOWS UPO TO 1 m PER HOUR, TOO FAST FOR JUST DIFFUSION OR CYTOPLASMIC STREAMING • THE FLOW IS BY A BULK FLOW (PRESSURE-FLOW) MECHANISM; BUILDUP OF PRESSURE AT THE SOURCE AND RELEASE OF PRESSRE AT THE SINK CAUSES SOURCE-TO-SINK FLOW • AT THE SOURCE END, PHLOEM LOADING CAUSES HIGH SOLUTE CONCENTRATIONS – WATER POTENTIAL DECREASES, SO WATER FLOWS INTO TUBES CREATING HYDROSTATIC PRESSURE • AT THE SINK END, THE WATER POTENTIAL IS LOWER OUTSIDE THE TUBE DUE TO THE UNLOADING OF SUGAR; OSMOTIC LOSS OF WATER RELEASES HYDROSTATIC PRESSURE – XYLEM VESSELS RECYCLE WATER FROM THE SINK TO THE SOURCE
PRESSURE FLOW IN A SIEVE TUBE