TRANSKRPSYON BASKILAYICILARI karyotlarda basklayclarn bakterilerdekine benzer ekilde alanlar

TRANSKRİPSİYON BASKILAYICILARI Ökaryotlarda baskılayıcıların bakterilerdekine benzer şekilde çalışanları çok enderdir. En yaygın biçimde olanı, baskılayıcıların (aktifleştiricilere benzer şekilde) nukleozom değiştiricileri bir araya toplamasıdır; bir araya toplanan enzimler kromatini sıkılaştırırlar veya transkripsiyon aygıtı tarafından tanınan grupları yok ederler. Örneğin, histonların kuyruklarındaki asetil gruplarının yok edilmesiyle transkripsiyonun baskılanması (histon deasetilazlar) Histon kuyruklarına metil grupları eklenerek transkripsiyonun baskılanması ( “sessizleştirme” şeklindeki baskılamanın da temeli).

Ökaryotlarda baskılamaya ait çeşitli yollar

Ökaryotlarda baskılamaya ait çeşitli yollar

SİNYAL İLETİMİ ve DÜZENLEYİCİLERİNİN KONTROLU TRANSKRİPSİYON Sinyaller genelde transkripsiyon düzenleyicileriyle sinyal iletimi yollarında ilişkiye girerler

GEN “SESSİZLEŞTİRİLMESİ” Genler özel bir çevresel sinyale yanıt verecek bir yerde bulunmadıkları zaman “sessizleşirler”. Sessizleştirilme yerleşim durumu etkisi Sessizleşme uzun DNA parçaları boyunca “yayılabilir”, başladığında bulunduğu yerden çok uzaktaki çok sayıda geni baskılar. Gen sessizleştirilmesi histonlarda ya da DNA’daki değişimlerle meydana gelir.

Sessizleşmenin en yaygın biçimi heterokromatin ile ilişkilidir. Heterokromatin özellikle, gen taşımayan, telomer ve sentromer bölgelerinde bulunur. Kromozomun başka yerlerindeki heterokromatin bölgelerde de genelde sessizleşmiş genler taşınır. (Memeli hücrelerinde, genomun ~%50’sinin heterokromatin durumunda )

Heterokromatik sessizleştirme Kromatinin yoğunluğunun histon kuyruklarını değiştiren enzimler tarafından değiştirilmesi. Yapıyı sıkıştırıcı etkiler DNA’nın tanınıp bağlanma derecesini etkiler ve transkripsiyonu etkiler. DNA metilazlar tarafından DNA’nın metillenmesi.

Histonların deasetillenmesi sessizleştirilmesi ve metillenmesiyle genlerin S. cerevisiae’de telomerdeki sessizleşme. Kromozomların ~1 -5 kb’lık kısmı telomerde kıvrılmış yoğunlaşmış durumdadır. Telomerdeki kromatin genomun (genlerin etkin anlatım yaptığı) diğer kısımlarından daha az asetillenmiş durumdadır. Telomerdeki sessizleşme düzenleyicilerini şifreleyen genler: SIR 2, 3 ve 4; (SIR, “silent information regulator”). Bu genlerin ürünleri olan proteinler sessiz kromatinle ilişkili bir kompleks oluştururlar; Sir 2 bir histon deasetilazdır.

Sessizleştirme kompleksi, telomerdeki tekrar dizileri tanıyıp DNA’ya bağlanan bir protein tarafından telomerde toplanır. Histon kuyruklarındaki asetil grupları yok edilir. Asetil grupları yok edilmiş histonlar, sessizleştirme kompleksi tarafından daha kuvvetli tanınır ve bölgesel deasetillenme kromatin boyunca yayılır ve yoğunlaşmış bir heterokromatin bölgesi yaratır. Yayılmanın telomerle (ve diğer heterokromatin bölgeleriyle) sınırlanması nasıl olur? Başka histon değişimleri (H 3 histon kuyruğunun metillenmesi) Sir 2 proteinlerinin bağlanmalarını engeller ve yayılmayı durdurur.

DNA’nın metillenmesiyle genlerin sessizleştirilmesi Bazı memeli genleri yakınlarındaki metillenmesiyle sessiz durumda tutulurlar. DNA dizilerinin DNA metillenmesi aynı zamanda heterokromatin bölgelerde görülür. Metillenmiş dizilerin genelde DNA’ya bağlanan proteinler tarafından tanınır; bu proteinler daha sonra kromatini değiştiren histon deasetilazları ve histon metilazları burada toplarlar. DNA’nın metillenmesi daha sonra heterokromatin oluşacak bölgeleri işaretleyebilir.

DNA’nın metillenmesiyle gen sessizleştirilmesi

Bir genin DNA metillenmesi ve histon değişimleriyle sesizleştirilmesi
 Temeli DNA metillenmesi Az sayıda bazı örneklerde, diploid bir hücrede bir genin")
Damgalama (“imprinting”) Temeli DNA metillenmesi Az sayıda bazı örneklerde, diploid bir hücrede bir genin (biri babadan diğeri anadan gelen) iki allelinden biri anlatım yaparken diğeri sessiz kalır.
H 19 ve")
İyi araştırılmış bir örnek insandaki (11. kromozomda birbirlerine çok yakın konumda bulunan)H 19 ve Igf 2 genleri: Bir hücrede H 19 geninin (anadan gelen) kopyası anlatım yaparken (babadan gelen) diğer kopyası sessiz kalır; Igf 2 geninde ise tersi olur. Bu genlerin farklı anlatım yapmalarında iş gören iki düzenleyici dizi : (H 19 geninin aşağısında) yükseltici ve (H 19 ve Igf 2 genleri arasında bulunan) yalıtıcı. Yükseltici neden sadece anaya ait kromozomdaki H 19 ve babaya ait kromozomdaki Igf 2 genini aktifleştirir? Bu özellik, yalıtıcının metillenme durumundan kaynaklanır.

Yükseltici anaya ait kromozomdaki Igf 2 genini aktifleştiremez; çünkü bu kromozomda yalıtıcı, yükselticideki aktifleştiricileri engelleyen bir proteine (CTCF) bağlı durumdadır. Babaya ait kromozomda ise yalıtıcı ve H 19 promotörü metillenmiş durumdadır. Bu durumda, transkripsiyon aygıtı promotöre ve CTFC de yalıtıcıya bağlanamaz. Sonuçta, yükseltici Igf 2 genini aktifleştirir.

TRANSKRİPSİYONUN BAŞLAMASINDAN SONRAKİ AŞAMALARDA GEN ANLATIMININ DÜZENLENMESİ Transkripsiyonda uzamanın düzenlenmesi Transkripsiyonun uzamasına yardım eden çeşitli aktifleştirici proteinler vardır. Bazı genlerde promotörün aşağı kısmındaki bazı diziler polimerazın transkripsiyonu başlattıktan hemen sonra duraklamasına yol açarlar. Bu genlerde, uzama faktörlerinin varlığı ya da yokluğu genin anlatım düzeyini büyük ölçüde etkiler.

Örneğin, Drosophila’da ısı şokuyla aktifleşen HSP 70 geni iki aktifleştirici tarafından kontrol edilir. GAGA dizisine bağlanan aktifleştirici başlama transkripsiyon aygıtının bir araya toplanmasında iş görür. için Diğer aktifleştiricinin (HSF) yokluğunda ise RNA polimeraz promotörün ~100 bç aşağısında duraklar. Isı şokuna yanıt olarak, HSF promotörde özel bir diziye bağlanır ve bir kinazı duraklamış aygıta getirir. Kinaz RNA polimerazın büyük alt biriminin C-ucu domenini fosforiller. Duraklamış olan enzim serbest kalır ve transkripsiyona devam eder.

Alternatif m. RNA kırpılmasının düzenlenmesi Öncül m. RNA’da alternatif kırpılmalar belli bir zamanda ya da belli bir hücrede hangi proteinin meydana geleceğinin seçiminin kontrolu. Alternatif kırpılmanın düzenlenmesinde belirleyici özellik: kırpılma yerinin kırpılmayı yapacak aygıta ve düzenleyici proteinlere olan ilgisinin değeri. Kuvvetli bir kırpılma yeri etkin ve sürekli bir kırpılmayı sağlar. Fakat kuvvetli kırpılma yeriyle üste gelen yerlere bağlanan kırpılma baskılayıcısı tarafından kırpılma engellenebilir ve kırpılma aygıtı dışarı atılır. Bazı örneklerde, kırpılma yerlerinin yakınında kırpılma yükselticileri bulunur. Bu diziler kırpılma aygıtını kırpılma yerinde toplayan düzenleyici proteinler tarafından tanınırlar. Düzenleyici proteinlerin RNA’ya bağlanma ve kırpılma aygıtına bağlanma domenleri ayrıdır.

GEN ANLATIMININ DÜZENLENMESİNDE ROL OYNAYAN RNA’LAR RNA’ların gen anlatımının düzenlenmesinde genel şekilde etkili olan ve farklı mekanizmayla meydana gelen rolleri de vardır. Enzimlerin etkisiyle yapılan bazı kısa RNA’lar onlarla homoloji gösteren genlerin anlatımını doğrudan baskılarlar [ RNA karışması, müdahalesi (“interference”) (RNAi) ] Baskılama, m. RNA’nın translasyon sırasındaki engellenmesi, m. RNA’nın yıkımı , m. RNA’nın anlatımını yöneten promotörün sessizleştirilmesi şeklinde olabilir. RNAi birçok organizmada özel genlerin anlatımını kapamakta teknik olarak kullanılmaktadır !!!
 C. elegans’ta hücrelere giren çift iplikli RNA’ların (ds. RNA)")
Çift iplikli RNA’lar (ds. RNA’lar) C. elegans’ta hücrelere giren çift iplikli RNA’ların (ds. RNA) onlara homolog (ya da çok benzer) olan genleri baskılayabildiği anlaşılmıştır. Benzer etkiler daha sonra başka organizmalarda da bulunmuştur. ds. RNA’lar Dicer olarak adlandırılan, RNAz. III tipi bir enzim tarafından tanınır ve parçalanırlar. Oluşan ürünler ~23 nukleotid uzunluğunda kısa çift iplikli parçalardır (si. RNA’lar). si. RNA’lar homolog bir genin anlatımını üç yolla engellerler: (1) m. RNA’sının yıkımını tetiklerler; (2) m. RNA’sının translasyonunu engellerler; (3) promotörde geni sessizleştirici kromatin değişimlerini tetiklerler.
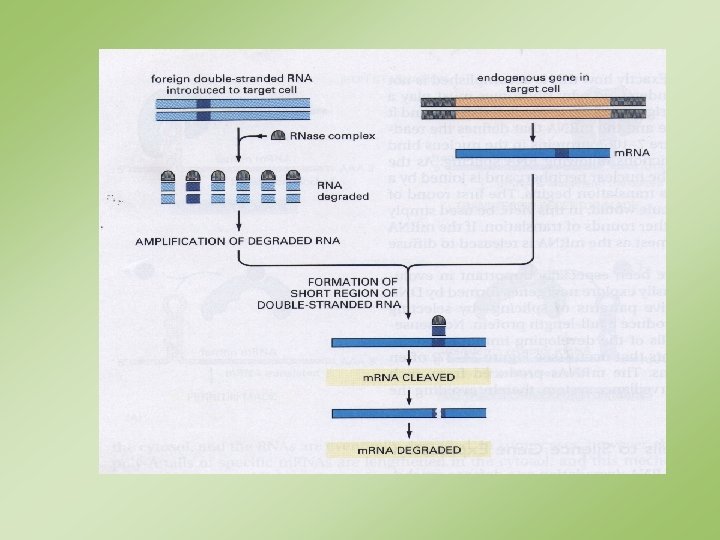
 oluşumu. Bu kompleks si.")
Yolların hepsinde genelde aynı mekanizma gereklidir: RISC (“RNA-induced silencing complex”) oluşumu. Bu kompleks si. RNA’lara ek olarak bazı proteinleri kapsar. Bir si. RNA meydana geldikten ve RISC ile birlik oluşturduktan sonra ATP’ye bağımlı şekilde denatürasyona uğrar. Tek iplikli RNA’nın ortaya çıkışı RISC kompleksini aktifleştirir. Aktifleşen kompleks si. RNA ile tamamlayıcı özellikteki bir m. RNA’ya yönelir. m. RNA’nın yıkımını yapar ya da translasyonunu engeller. Seçilen yol, si. RNA ile hedef m. RNA arasındaki eşleşmenin nasıl olduğuna bağlıdır: eşleşme çok iyi değilse yanıt translasyonun engellenmesidir; tamamlayıcılık tam ise RNA’nın yıkımıdır; yıkımdan RISC içindeki bir nukleaz aktivitesi sorumludur.

Ayrıca, RISC kompleksi bir si. RNA tarafından nukleus içindeki si. RNA ile tamamlayıcılık ilişkisi gösteren bölgelere yönlendirilir. Kompleks o bölgelerdeki genlerin promotörlerinin etrafındaki kromatini değiştirecek diğer proteinleri oraya toplar. Bu değişim o genlerde (sessizleşmesine) yol açar. transkripsiyon yapmamasına
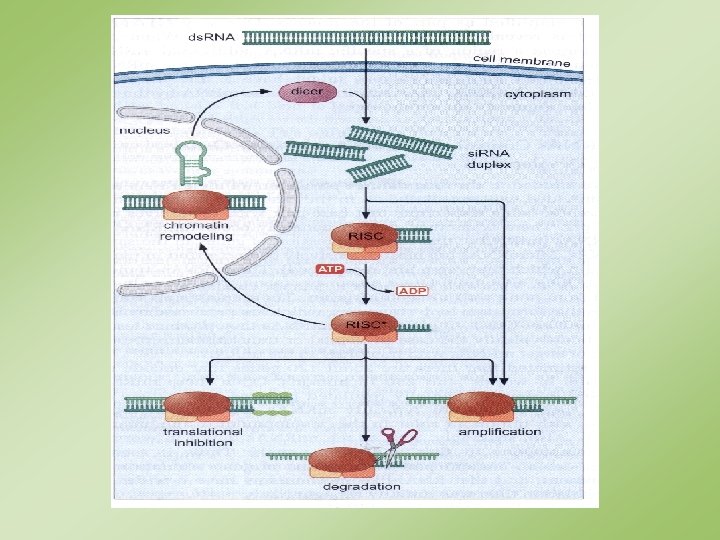

RNAi sessizleştirmesinin çok önemli bir özelliği: ds. RNA’ların çok küçük miktarları hedef genlerin tamamen kapatılması için yeterlidir. Bu kuvvetli bir etkinin mekanizması henüz açık değil. Belki, RNA’ya bağımlı RNA polimeraz aktivitesi etkili olabilir. Böyle bir enzimin işe karışması engelleyici “sinyal”in çoğalmasına yol açar. Bir si. RNA özel bir m. RNA’nın bir bölgesini hedeflediği zaman, genelde aynı m. RNA’daki komşu bölgeleri hedefleyen ek si. RNA’lar üretilir. RNA’ya bağımlı RNA polimeraz orijinal si. RNA tarafından m. RNA’da toplandıktan sonra bu ek RNA’ların üretiminde rol oynayabilir.
 Doğal olarak meydana gelen iç kaynaklı küçük RNA sınıfıdır. mi.")
Mikro RNA’lar (mi. RNA’lar) Doğal olarak meydana gelen iç kaynaklı küçük RNA sınıfıdır. mi. RNA’lar bitkilerde ve solucanlarda iyi tanımlanmıştır. mi. RNA’lar genelde 21 veya 22 nukleotid boyundadır. Kökenleri: protein şifrelemeyen mi. RNA genlerinin veya yapısal genlerin RNA polimeraz II tarafından kopyalanma ürünlerindeki (70 -90 nukleotid boyundaki) intron veya ekson bölgeleri mi. RNA öncüllerini şifreleyen solucanlarda ~120, insanlarda ~250 gen bulunur. Öncül mi. RNA’lar ardışık önce bir RNaz. III (Drosha) tarafından işlenir ve sap-ilmek biçiminde ve 3’ ucunda 2 nt çıkıntı bulunan yapılar oluşur (~70 -80 nt); bu moleküller bir “exportin” tarafından nukleustan çıkarılır ve sitoplazmada, Dicer tarafından işlenir aktif mi. RNA’lar

mi. RNA’lar genlerin baskılanmasını si. RNA’lara benzer şekilde yönetirler; ya homolog m. RNA’ların yıkımına (bitkilerde), ya da translasyonunun engellenmesine (solucanda) yol açarlar. mi. RNA’lar genelde gelişim sırasındaki gen anlatımı düzenlemesinde rol oynarlar ve hedefleri organizmanın gelişiminde önemli rolleri olan düzenleyici proteinlerin m. RNA’larıdır.
- Slides: 27