Transcriptional Gene Regulation in Eukaryotes Overview 4 Gene












 Promoter 4 Proximal cis-regulatory module")

")






")













 – DNA binding (helix) & dimerization – Class A: ubiquitouslyh expressed")






; by RNA processing")







")
 • Twist (HLH) • a HLH • Snail (-)")

 4 Secreted embryonic gut")

 4")


")
, histone deacetylase (HDAC)")
.")





 and")







- Slides: 82
Transcriptional Gene Regulation in Eukaryotes
Overview 4 Gene expression 4 Transcription Regulation of eukaryotic transcription Influence of chromatin structure Oncogenes Techniques 4 4
Orphanides, Cell 2002
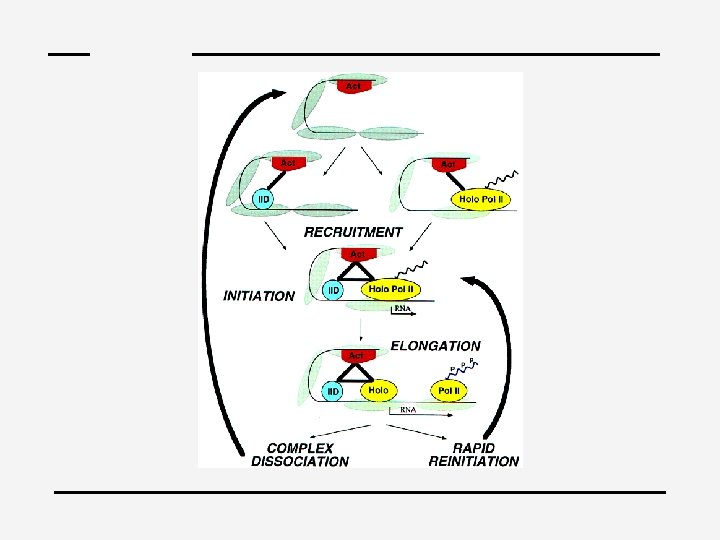
4 Control Activation of gene structure of gene expression at any stage: Initiation of transcription
What is a gene? 4 “The entire nucleic acid sequence that is necessary for the synthesis of a functional polypeptide or RNA molecule”
Overview Gene expression 4 Transcription 4 Regulation of eukaryotic transcription 4 Influence of chromatin structure 4 Oncogenes 4 Techniques 4
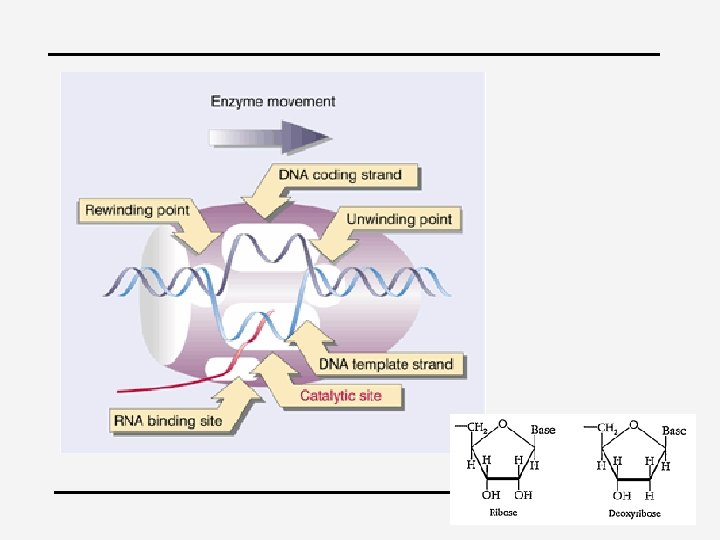
Transcription 4 Initiation, elongation, termination 4 Catalyzed by RNA polymerase – “Transcription bubble”: DNA transiently separated into single strands – One strand is used as a template – Unwinding point & rewinding point – Rate 40 nucleotides/second at 37 for bacteria 4 RNA polymerase – Many subunits: catalytic site, CTD with (YSPTSPS)n – pol I, pol III
Initiation
Bacteria
4 Molecular details of gene expression control in bacteria: lac operon E. coli (Jacob & Monod 1960 s)
Bacterial enhancer Glutamine synthase
Eukaryotes
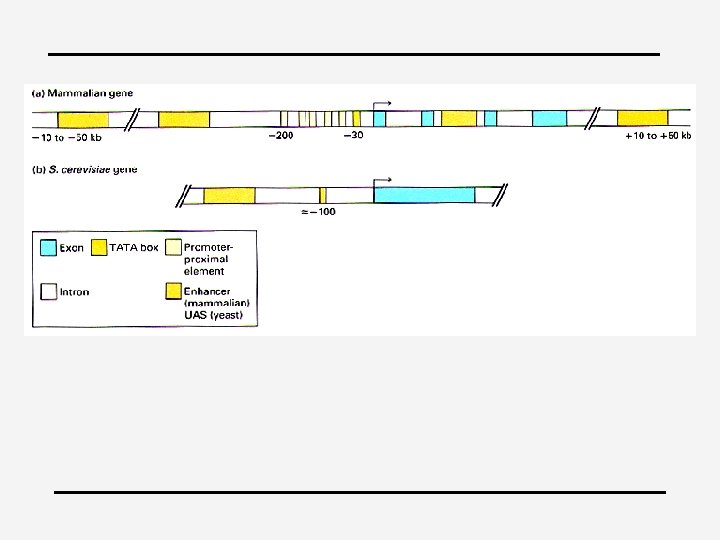
4 Basal transcription apparatus (general factors & RNA polymerase) Promoter 4 Proximal cis-regulatory module Enhancer 4 Distal cis-regulatory modules 4 Modules = discrete DNA elements that contain specific sequence motifs with which DNA binding proteins interact and transmit molecular signals to genes
BTA – General factors: TFIIx – Mechanics of initiating RNA synthesis at all promoters – Determines location of transcription startpoint – Complex with RNA polymerase – TATA • ~ 25 bp upstream • 8 bp consensus of A • T pairs • Tends to be surrounded by G • C rich regions • TBP, 11 TAFs : TFIID (~800 k. D)
TATA – TATA-less promoters: Inr • Inr Py 2 CAPy 5 (-3 to +5) TATA – Promoter-prediction: TATA-box, C-G enrichment Promoter 4 3 – 50% Eponine 6 12 132 23 19 Inspector Cp. G islands
Promoter-proximal region 4 Efficiency and specificity of transcription depend on binding of transcription factors
Promoter recognition 4 Function = to be recognized by proteins; so differs from exon, … 4 Any essential nucleotide sequence should be conserved – Some variation is permitted – When is it sufficiently conserved? – Idealized sequence with base most often present: consensus sequence by aligning all known examples – Only conservation of very short sequences; 60 bp associated with RNA pol lack conservation
Variety of elements can contribute, none is essential for all promoters (mix & match principle) 4 CAAT box ~ -80 bp GGCCAATCT – increases promoter strength – Bound by CTF/NF 1 family, CP 1 & CP 2, C/EBP, ACF 4 GC box – SP 1 GGGCGG 4 Octamer (8 bp) ATTTGCAT – Bound by Oct 1 (ubiquitous): activates histon H 2 B – Bound by Oct 2 (lymphoid cells): Ig kappa light chain – context is important
4 Modular nature of the promoter: – Equivalent regions can be exchanged – Main purpose = to bring the factors they bind into the vicinity of the initiation complex – Protein-protein interactions determine the efficiency of the initiation reaction 4 Sequence elements influence the frequency of initiation 4 Repression of transcription: – Generally by influencing chromatin structure – By repressors, e. g. Dr 1/DRAP 1 binds to TBP and CAAT displacement protein (CDP)
Modules 4 Enhancers, silencers 4 5’ region, distal
Modules 4 4 4 50 bp to 1. 5 kbp in size 4 -8 TFs (often multiple sites); higher density of regulatory elements than in the promoter Many elements are common elements in promoters, e. g. AP 1 and the octamer Can stimulate any promoter placed in its vicinity Can function anywhere (cfr -globin: 200 fold in vivo) ; Position relative to promoter can vary substantially; can function in either orientation
Example: TTR Binding sites for activators that control transcription of the mouse transthyretin (TTR) promoter in hepatocytes. HNF = hepatocyte nuclear factor. [See R. Costa et al. , 1989, Mol. Cell Biol. 9: 1415; K. Xanthopoulus et al. , 1989, Proc. Nat’l. Acad. Sci. USA 86: 4117. ]
Example: muscle specific modules Creatine kinase, myosin light chain, skeletal actin, myosin heavy chain
Example: -globin Model for the control of the human b-globin gene. Some of the gene regulatory proteins shown, such as CP 1, are found in many types of cells, while others, such as GATA-1, are present in only a few types of cells including red blood cells and therefore are thought to contribute to the cell-type specificity of b globin gene expression. (Adapted from B. Emerson, In Gene Expression: General and Cell-Type-Specific [M. Karin, ed. ], pp. 116 -161. Boston: Birkhauser, 1993. )
How?
4 Current view: – same sort of interaction with basal apparatus as the proximal promoter module – Increase the concentration of transcription factors in the vicinity of the promoter 4 Intervening DNA: extruded as a large “loop” 4 Generality: not yet clear (what proportion of promoters require an enhancer? )
Cooperative assembly Four activators enriched in hepatocytes plus the ubiquitous AP 1 factor bind to sites in the hepatocytespecific enhancer and promoter-proximal region of the TTR gene. The activation domains of the bound activators interact extensively with co -activators, TAF subunits of TFIID, Srb/Mediator proteins, and general transcription factors, resulting in looping of the DNA and formation of a stable activated initiation complex.
Limited knowledge 4 Experimentally verified binding sites 4 Experimentally verified “composite elements” or CE’s – GR site + AP-1 in proliferin promoter – Synergistic: result in non-additively high level – Antagonistic: overlapping sites, masking an activation domain, … – Direct or through coactivator 4 Few modules characterized that have multiple elements, some in developmental biology
Side-track: Transcription factors 4 5% of our proteins 4 Activities controlled in regulatory pathways 4 Independent domains responsible for activities: – Recognition of specific target sequences – Binding to other components of the transcription apparatus – E. g. yeast GAL 4
4 Protein-DNA interactions – Proteins with high affinity for a specific sequence also possess a low affinity for any (random) DNA sequence – E. g. Lac repressor E. coli: Free: bound = 10 -4 – High-affinity site competes with the large number of low-affinity sites; repressor binds 107 times better to operator DNA (bound 96% of time for 10 molecules/cell)
How the different base pairs in DNA can be recognized from their edges without the need to open the double helix.
The binding of a gene regulatory protein to the major groove of DNA. Typically, a protein-DNA interface consists of 10 to 20 such contacts, involving different amino acids, each contributing to the binding energy of the protein-DNA interaction.
4 Zinc finger motif – Common motif in DNA binding, e. g. SP 1 has 3 (A) The structure of a fragment of a mouse gene regulatory protein bound to a specific DNA site. This protein recognizes DNA using three zinc fingers of the Cys-His-His type arranged as direct repeats. (B) The three fingers have similar amino acid sequences and contact the DNA in similar ways. In both (A) and (B) the zinc atom in each finger is represented by a small sphere. (Adapted from N. Pavletich and C. Pabo, Science 252: 810 -817, 1991. © 1991 the AAAS. )
4 Helix-Turn-Helix All of the proteins bind DNA as dimers in which the two copies of the recognition helix (red cylinder) are separated by exactly one turn of the DNA helix (3. 4 nm). The second helix of the helix-turn-helix motif is colored blue. The lambda repressor and cro proteins control bacteriophage lambda gene expression, and the tryptophan repressor and the catabolite activator protein (CAP) control the expression of sets of E. coli genes.
4 Homeodomains – Related to helix-turn-helix bacterial repressors – Homeobox = 60 AA residues – E. g. en, eve, Hox, Oct-1, Oct-2 (Oct also have Pou domain next to homeodomain) The homeodomain is folded into three alfa helices, which are packed tightly together by hydrophobic interactions (A). The part containing helix 2 and 3 closely resembles the helix-turn-helix motif , with the recognition helix (red) making important contacts with the major groove (B). The Asn of helix 3, for example, contacts an adenine. Nucleotide pairs are also contacted in the minor groove by a flexible arm attached to helix 1. The homeodomain shown here is from a yeast gene regulatory protein, but it is nearly identical to two homeodomains from Drosophila, which interact with DNA in a similar fashion. (Adapted from C. Wolberger et al. , Cell 67: 517 -528, 1991. © Cell Press. )
4 Helix-loop-helix (HLH) – DNA binding (helix) & dimerization – Class A: ubiquitouslyh expressed proteins, e. g. E 12/E 47 – Class B: tissue-specific expression, e. g. Myo. D, myogenin, Myf-5 – Myc proteins (separate class) 4 Leucine zippers fig 21. 15 – Dimerization motif – E. g. Jun+Fos = AP 1 – Gcn 4 ->
4 Steroid receptors – Independent domains: DNA binding, hormone binding, and dimerization Cortisol - glucocorticoid receptor (GR). Retinoic acid - retinoic acid A receptor (RAR). Thyroxine - thyroid hormone receptor (TR).
Figure 1 Genome-wide comparison of transcriptional activator families in eukaryotes. The relative sizes of transcriptional activator families among Homo sapiens, D. melanogaster, C. elegans and S. cerevisiae are indicated, derived from an analysis of eukaryotic proteomes using the INTERPRO database, which incorporates Pfam, PRINTS and Prosite. The transcription factors families shown are the largest of their category out of the 1, 502 human protein families listed by the IPI.
Alternative model
Transcription factories 4 cfr. replication factories 4 Active RNA polymerases are concentrated in discrete 'factories' where they work together on many different templates 4 Complexes for transcription and RNA processing are likely to be immobile structures within the gel-like nucleoplasm (Burns et al, 2001; Kimura et al, 1999) 4 Transcriptional interference: phenomenon where transcription of one gene prevents transcription of an adjacent gene. Discovery: Cells were transfected with a retroviral vector encoding resistance to neomycin and azaguanine, and clones harboring a single copy of the vector selected. Expression of the 3' gene was suppressed when selection required expression of the 5' gene, and vice versa. In addition, hardly any cells grew in both neomycin and azaguanine
• Enhancers Cook, 1999 • dynamic equilibrium • enhancing the probability of the key transcription cycle interactions • Element 5’ or 3’ doesn’t matter! (Science)
Recap: evolution of understanding of eukaryotic transcription Lemon and Tjian, Genes Dev. 14: 2551 -2569 (2000)
Termination 4 Bacteria 4 Eukaryotes (? ); by RNA processing
Overview 4 Gene expression 4 Eukaryotic transcription 4 Regulation of eukaryotic transcription Influence of chromatin structure 4 Oncogenes 4 Techniques 4
Activate/inactivate a TF 4 Transport through nuclear pores from cytoplasm to nucleus (e. g. masking NLS, nuclear localization signal, can regulate this transport) 4 Link to Ubiquitin protease system – Rapid turnover of promoter bound TF: resets signaling pathway: cell can continuously monitor its environment 4 Tissue-specific synthesis – Development, e. g. homeodomain proteins 4 Modification – Phosphorylation, acetylation, methylation – E. g. , AP 1 (= Jun+Fos) active form by phosphorylation – E. g. , p 53 acetylated (modulates interactions with coactivator and repressor proteins
4 Ligand binding – E. g. Steroid receptors – Influence: localization or DNA-binding ability 4 Cleavage 4 Inhibitor release – E. g. NF- B + I- B (release in B lymphocytes) 4 Change of partner (active partner displaces inactive partner)
Examples: GATA-1 CAP Ntr. C Pathways… Adenovirus E 1 A + CBP/p 300 NF-KB/ glucocorticoid receptor
2 1
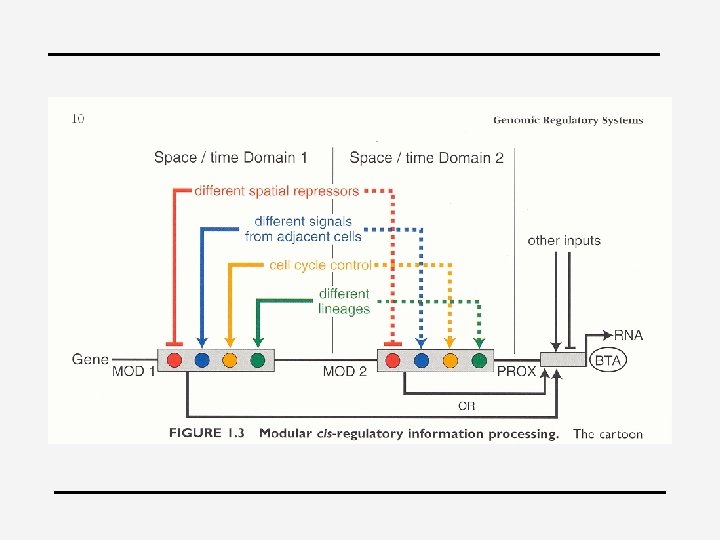
4 Level 1 = active/inactive factor 4 Level 2 = cooperation of multiple factors within a module (all present and active, and all repressors inactive or absent) 4 Level 3 = multiple autonomous modules per gene – Each module can independently activate the gene – Each has a specific function (e. g. activation in certain cell type or at particular stage in dvl) – different circuits of regulation, e. g. metallothionein gene (MT): heavy metals and steroids, fig 21. 1 – Gene can respond to multiple signaling pathways – Facilitates fine-tuning of transcript levels
4 Combinatorial and context dependent regulation of transcription – one factor can induce transcription of one gene while repressing that of another
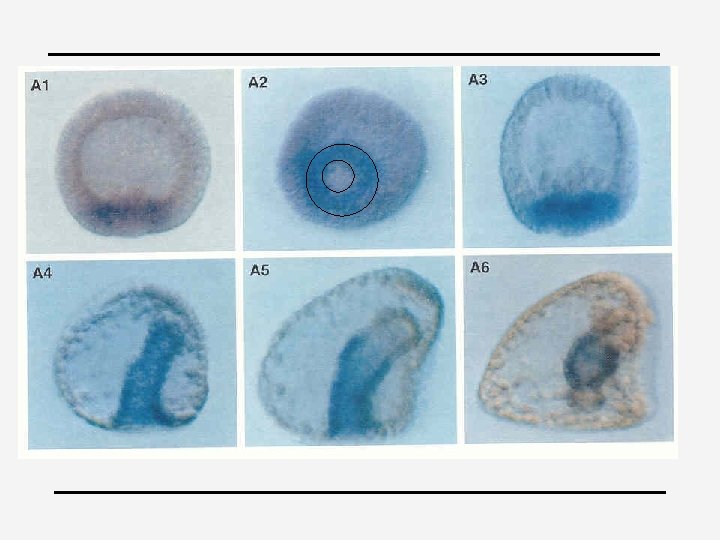
Example: eve Experiment demonstrating the modular construction of the eve gene regulatory region. (A) A 480 -nucleotide-pair piece of the eve regulatory region was removed and inserted upstream of a test promoter that directs the synthesis of the enzyme galactosidase (the product of the E. coli lac. Z gene). (B) When this artificial construct was reintroduced into the genome of Drosophila embryos, the embryos expressed -galactosidase (detectable by histo-chemical staining) precisely in the position of the second of the seven eve stripes (C). (Metamerization) +
rho • Dorsal (Dl) • Twist (HLH) • a HLH • Snail (-)
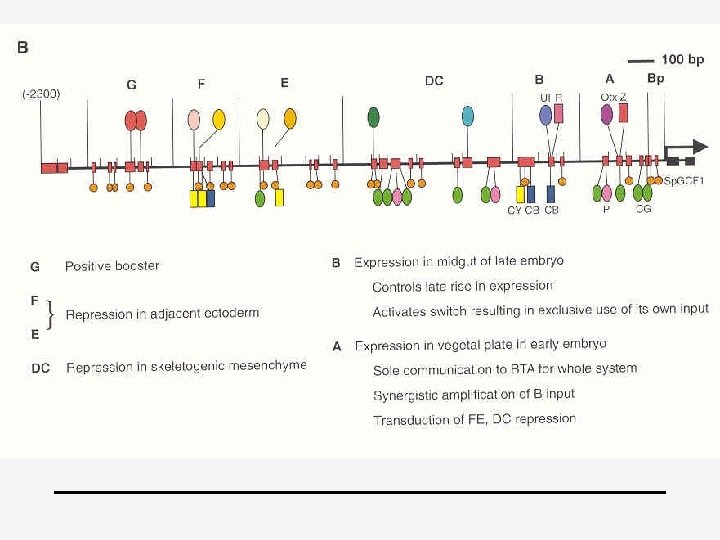
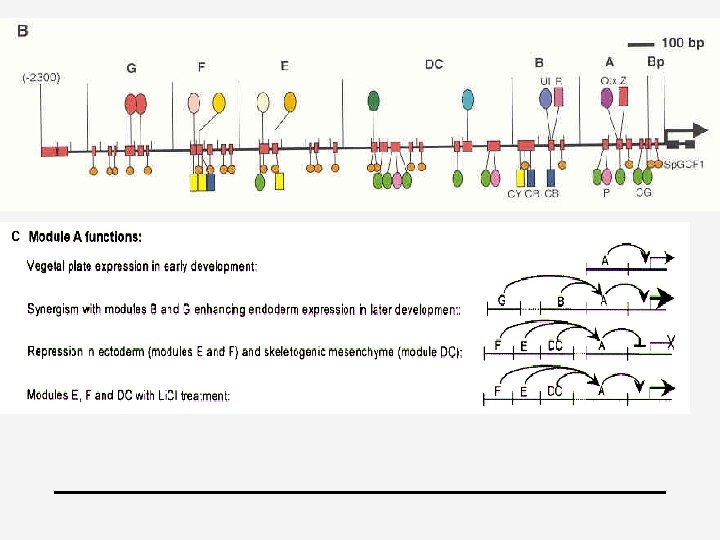
Principles for specification 4 4 4 1. cis-regulatory transformation of input patterns into spatial domains of differential gene expression 2. Always assemblages of diverse target sites because multiple inputs are required 3. Output=novel with respect to any one of the incident inputs + more precise in space and time => “information processing” 4. Every specific type of interaction that can be detected in vitro is fundamentally significant (it is unlikely that highly specific site clusters, which are of improbable random occurrence would have no function) 5. Negative & positive inputs (Davidson, 2001)
4 Cis-regulatory logic device 4 endo 16 of Strongylocentrotus (zee-egel) 4 Secreted embryonic gut protein 4 “hardwired biological computational device”
Overview Gene expression 4 Initiation of transcription 4 Regulation of transcription 4 4 Influence of chromatin structure Oncogenes 4 Techniques 4
Chromatin Eukaryotic genomes are packaged with chromatin proteins 4 Heterochromatin (highly condensed, untranscribed) 4 Euchromatin (more accessible, transcribed) 4 Each cell: unique pattern of heterochromatin and euchromatin 4
Nucleosomes • 146 bp • H 2 A, H 2 B, H 3, H 4 Workman and Kingston Ann. Rev. Biochem. 67: 545 (1998)
Chicken and egg scenario 4 TF binding requires chromatin decompaction by certain factors but the latter also need to interact with DNA 4 Solution: probably some TFs can bind to their recognition sequences even when they are packaged (e. g. glucocorticoid receptor: only contacts DNA on one side NF 1 surrounds double helix)
Modify chromatin structure 1. ATP-dependent remodeling Narlikar, 2002 (Cell)
2. Histone-modifying complexes • Phosphorylation, methylation, acetylation • Histone acetyltransferase (HAT), histone deacetylase (HDAC) • How do they impact the structure of the template and the ability of the transcription machinery to function? • lowered positive charge on acetylated N termini, lowered stability of interaction with DNA • Disrupting internucleosomal interactions • Recruiting additional TFs • A lot of combinatorial possibilities: histon code?
Jenuwein T, Allis CD. Science 293: 1074 -80 (2001).
Model of the protein interactions and functions of the Myc/Max/Mad transcription network. Grandori C, Cowley SM, James LP, Eisenman RN. Annu Rev Cell Dev Biol. 2000; 16: 65399 Myc-Max and Mad-Max (along with Mnt-Max and Mga-Max) complexes bind to DNA to E-boxes. Binding can be affected by the context, sequence, cooperativity, and location of the E-boxes. Myc-Max heterodimers activate transcription by recruiting HAT's via TRRAP. This leads to the acetylation of histone tails and the opening of local chromatin structure. Additionally, Myc-Max appears to repress transcription through Inr elements via an undefined mechanism. As a result of these activities at target genes, Myc affects proliferation, cell cycle, growth, immortalization, and apoptosis. When deregulated, Myc cooperates with other oncogenes to cause a variety of cancers. Mad-Max and Mnt-Max heterodimers repress transcription by recruiting HDAC's via m. Sin 3 A. This leads to the deacetylation of histone tails and the closing of local chromatin structure. As a result of target gene repression, Mad causes an increased cell doubling time, growth arrest, and the maintenance of differentiation.
Cytosine methylation 4 m. CG often in inactive vertebrate genes 4 After replication of methylated DNA, methyl groups are added to daughter strands 4 Cp. G islands 4 imprinting
Imprinting Imprinted genes are genes whose expression is determined by the parent that contributed them. 4 Imprinted genes violate the usual rule of inheritance that both alleles in a heterozygote are equally expressed. 4 Examples of the usual rule: 4 – If a child inherits the gene for blood group A from either parent and the gene for group B from the other parent, the child's blood group will be AB. – If a child inherits the gene encoding hemoglobin A from either parent and the gene encoding hemoglobin S from the other parent, the child's red blood cells will contain roughly equal amounts of the two types of hemoglobin. 4 But there a few exceptions to this rule. A small number of genes in mammals (~50 of them at the most recent count) have been found to be imprinted. Because most imprinted genes are repressed, either – the maternal (inherited from the mother) allele is expressed exclusively because the paternal (inherited from the father) allele is imprinted or – vice-versa.
Link between DNA methylation and histone deacetylation
Chromatin compartmentalization 4 Heterochromatin: gene poor, inaccessible, perifery, transcr. inactive 4 Euchromatin: gene rich, transc. active
4 Consistent correlation between gene silencing (e. g. in B en T lymphocytes) and presence in heterochromatin regions – LCR, enhancers, insulators: act by maintaining endogenous loci in a chromatin compartment that is either transcr. permissive or nonpermissive?
Position variegation Position effects can be observed for the Drosophila white gene. Wild-type flies with a normal white gene have red eyes. If the white gene is inactivated by mutation, the eyes become white (hence the name of the gene). In flies with a chromosomal inversion that moves the white gene near a heterochromatic region, the eyes are mottled, with red and white patches. The white patches represent cells where the white gene is silenced and red patches represent cells that express the white gene. (After L. L. Sandell and V. A. Zakian, Trends Cell Biol. 2: 10 -14, 1992. )
Overview Gene expression 4 Initiation of transcription 4 Regulation of transcription 4 Alteration of chromatin structure during transcription 4 4 Oncogenes 4 Techniques
The development and metastasis of human colorectal cancer and its genetic basis. A mutation in the APC tumor-suppressor gene in a single epithelial cell causes the cell to divide, although surrounding cells do not, forming a mass of localized benign tumor cells called a polyp. Subsequent mutations leading to expression of a constitutively active Ras protein and loss of two tumor-suppressor genes, DCC and p 53, generates a malignant cell carrying all four mutations; this cell continues to divide and the progeny invade the basal lamina that surrounds the tissue. Some tumor cells spread into blood vessels that will distribute them to other sites in the body. Additional mutations cause exit of the tumor cells from the blood vessels and growth at distant sites; a patient with such a tumor is said to have cancer. [Adapted from B. Vogelstein and K. Kinzler, 1993, Trends Genet. 9: 101. ]
Overview 4 Gene expression Initiation of transcription Regulation of transcription Alteration of chromatin structure during transcription Oncogenes 4 Techniques 4 4
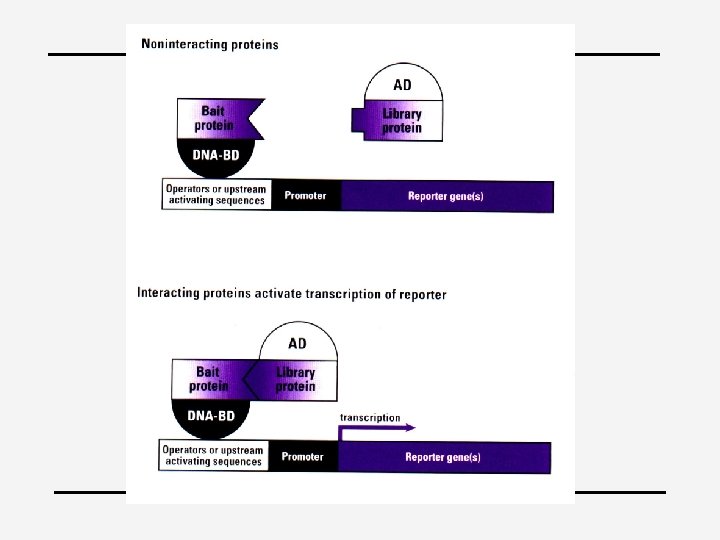
Techniques 4 Mutational 4 EMSA analysis electorphoretic mobility shift assay 4 Microarrays: level of m. RNA 4 Ch. IP: Chromatin Immuno. Precipitation 4 Yeast 2 Hybrid 4 Sequence analysis, phylogenetic footprinting
EMSA * Protein-DNA complex * Free DNA probe
Sources 4 B Lewin, Genes VII 4 Lodish et al. Molecular Cell Biology 4 EH Davidson: Genomic Regulatory Systems 4 Alberts et al. Essential Cell Biology 4 EM Blackwood & JT Kadonaga: Going the distance: a current view of enhancer action 4 Cell, February 22, 2002: 108 (4) "Reviews on Gene Expression"