Struktura membrn Penos ltek pes membrny Vznam bunnch

Struktura membrán Přenos látek přes membrány

Význam buněčných membrán • Plasmatická membrána odděluje buněčné složky od okolního prostředí • Umožňuje organelám vykonávat specializované funkce udržováním obsahu organel odděleně od zbytku buňky (kompartmentalizace) • Zajišťuje rozhraní pro vytváření elektrochemických gradientů, které jsou nutné pro syntézu ATP a vytváření nervových vzruchů • Reguluje transport živin do buňky a odpadních látek ven z buňky • Zachycuje signály z mimobuněčného prostředí • Interaguje s jinými buňkami nebo s extracelulární matrix (u mnohobuněčných organismů)

Amfipatické lipidy Proteiny Nekovalentní interakce
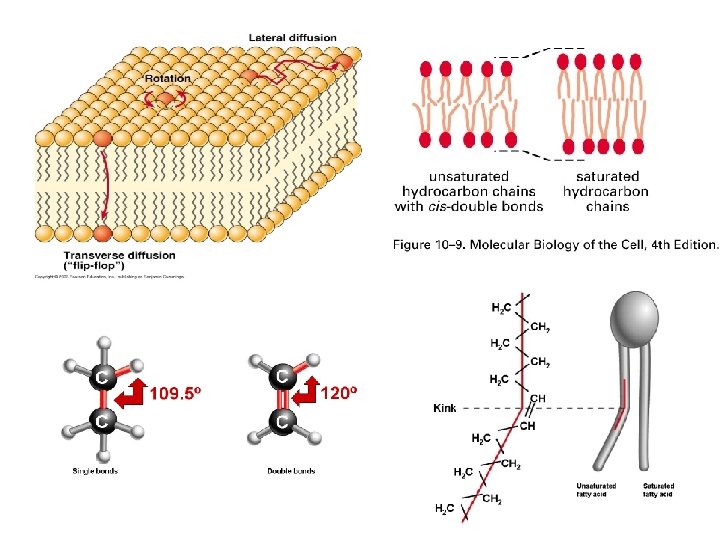
 Uhlovodíkový konec odvozený z mastných kyselin Nenasycený uhlovodíkový")
Polární hlavička Nepolární konec fosfatidylcholin (fosfolipid) Uhlovodíkový konec odvozený z mastných kyselin Nenasycený uhlovodíkový řetězec Nasycený uhlovodíkový řetězec

Mastné kyseliny * * *

Tři typy membránových lipidů

Fosfolipidy
 prostředím a svými nepolárními částmi.")
Amfipatické molekuly minimalizují interakce mezi vodním (polárním) prostředím a svými nepolárními částmi.

Cholesterol • Cholesterol snižuje fluiditu membrány protože omezuje pohyb uhlovodíkových řetězců. Důležité chemické vlastnosti cholesterolu: • Hydroxylová skupina představuje polární hlavičku. • OH je navázána na pevný cholesterolový zbytek. • Má jeden uhlovodíkový konec.

Fluidita membrány

Fluidita plasmatické membrány
")
FRAP (Fluorescence Recovery After Photobleaching)
")
FLIP (Fluorescence Loss In Photobleaching)
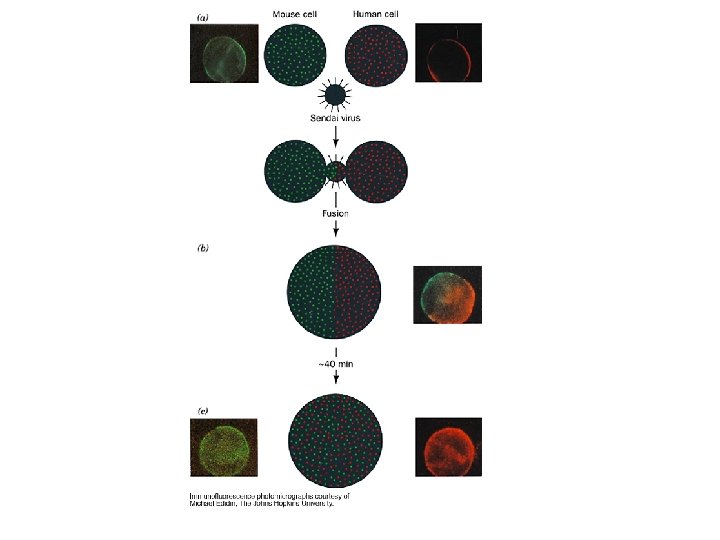

Růst membrán

Polarizace membrán Fosfatidylcholin Sfingomyelin Glykolipidy Cholesterol Fosfatidylserin Fosfatidylethanolamin Fosfatidylinositol

Polarizace membrán

Glykolipidy
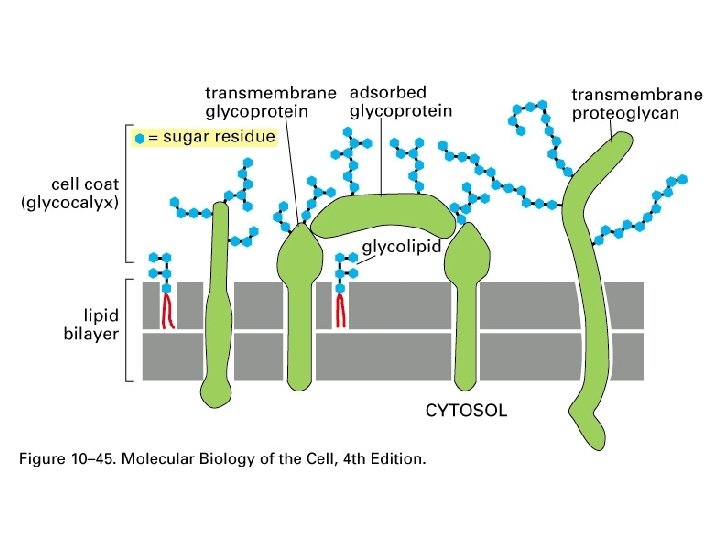
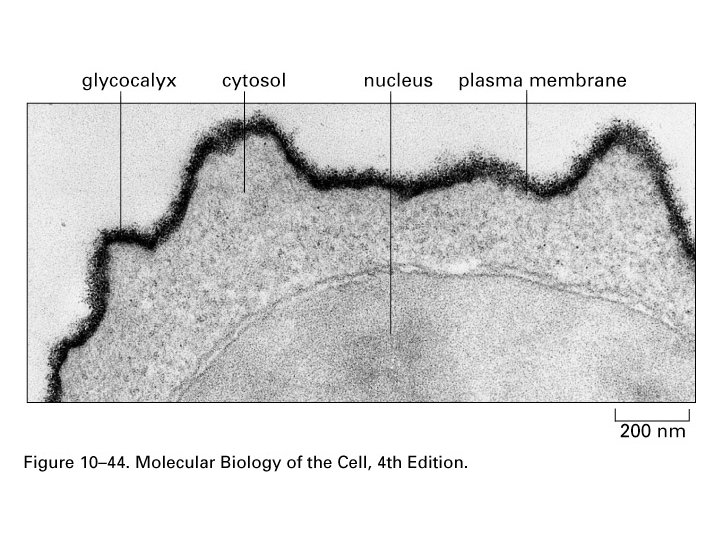

Polarita cytoplasmatické membrány

Membránové proteiny Transport Komunikace mezi buňkami Enzymatické aktivity

Typy membránových proteinů

alpha Helix

Typy aminokyselin

Transmembránová alfa šroubovice Tloušťka hydrofobní části lipidické membrány je 3 nm. Jedna otáčka šroubovije měří 0. 54 nm and je tvořena 3. 5 a. a. Takže transmembránová doména tvořená alfa-šroubovicí je dlouhá přibližně 20 aminokyselin.

Hydropatický graf
")
Hydropathy plot (Hydropatický graf)

Glycophorin Další vlastnosti mnoha alfa helikálních transmembránových proteinů: • SH skupiny na cytoplasmatické straně membrány jsou redukované a na vnější straně jsou oxidované na S-S. • Cukry jsou kovalentně navázány na určité aminokyseliny na vnější straně plasmatické membrány (glykosylace).
 a je složen z antiparalelních")
* Protein 3 je znám jako beta-soudek (beta barrel) a je složen z antiparalelních beta skládaných listů stočených do soudkovitého tvaru. • Mnohem vzácnější než alfa-šroubovicové transmembránové domény – omezen na vnější membrány bakterií, chloroplastů a mitochondrií (Porin). • Polární aminokyselinové postranní řetězce jsou umístěny uvnitř soudku a nepolární aminokyselinové řetězce se nacházejí na vnější straně.

beta-skládaný list

beta barrels transporter receptor enzyme channel

Buněčný kortex

Přenos látek přes membrány

Propustnost plasmatické membrány
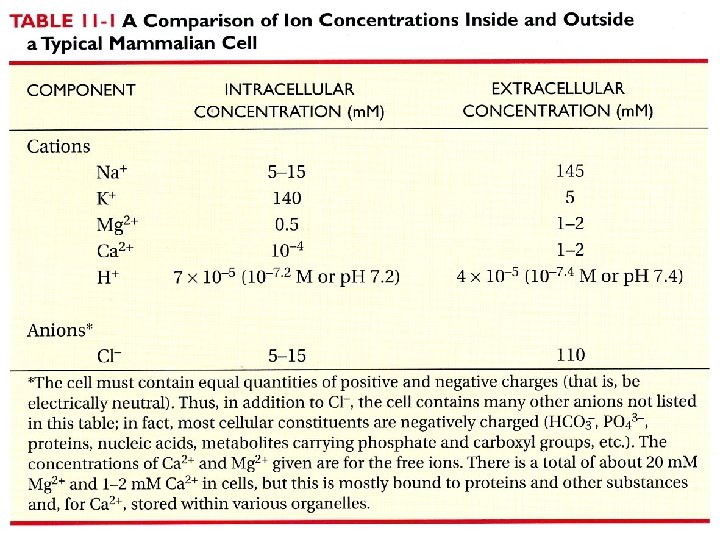

Transport molekul, které nejsou propustné přes plasmatickou membránu je zajišťován dvěmi hlavními třídami membránových transportních proteinů. Konformační změna “přenese” molekulu přes membránu. Vodní pór umožňuje průchod molekul přes membránu.

Čtyři cesty molekul a iontů přes membránu
 Přenašečový protein naváže transportovanou molekulu na jedné straně membrány, projde")
Přenašečové proteiny (Carrier proteins) Přenašečový protein naváže transportovanou molekulu na jedné straně membrány, projde konformační změnou a uvolní molekulu na druhé straně membrány.

Přenašečové proteiny jsou podobné enzymům Enzym Vazebná místa pro substrát Chemicky modifikuje substrát Rychlost reakce je saturována při vysoké koncentraci substrátu Přenašečový protein Vazebná místa pro transportované molekuly Transportuje molekuly Rychlost transportu je saturována při vysoké koncentraci přenášené látky

Spojením konformační změny se zdrojem energie mohou přenašečové proteiny provádět aktivní transport.

V závislosti na tom kolik různých molekul je transportováno a kterým směrem, rozlišujeme tři základní typy transportu: uniport, symport a antiport.
 v plasmatické membráně je antiporter, který provádí aktivní transport. Tento")
Na+-K+ pumpa (Na+-K+ ATPase) v plasmatické membráně je antiporter, který provádí aktivní transport. Tento protein vytváří koncentrační gradient Na+ a K+ iontů.

Na+ gradient vytvořený Na+ - K+ ATPázou pohání transport glukózy do buňky pomocí Na+ -poháněného glukózového symporteru. Energeticky příznivý pohyb Na+ po svém electrochemickém gradientu je spřažen s energeticky nepříznivým transportem glukózy proti svému koncentračnímu gradientu.

Iontové kanály a Membránový potenciál

Iontové kanály jsou iontově selektivní a uzavíratelné boční pohled R R R horní pohled – každý kruh představuje alfa-helix a Rskupiny jsou aminokyselinové postranní řetězce, které uvnitř kanálu vytvářejí tzv. selekční filtr (selectivity filter). R R R

Selectivita sodného kanálu

Jak může K+ kanál rozlišit mezi K+ and Na+ když oba mají stejný náboj a Na+ je menší K+? Odpověď: Při průchodu iontu přes selekční filtr, se iont musí zbavit vodního obalu. Molekuly kyslíku z karboxylových skupin mají částečný záporný náboj a mohou nahradit molekuly vody u K+, ale Na+ je příliš malý. Tudíž Na+ zůstává přednostně asociován s vodou a hydratovaný iont je příliš veliký pro průchod selekčním filtrem.

Iontové kanály oscilují mezi otevřeným a uzavřeným stavem za účelem regulace toku iontů. • Napěťově (elektricky) ovládané kanály reagují na membránový potenciál. • Chemicky ovládané kanály reagují na navázání malých molekul = ligandy. • Mechanicky ovládané kanály reagují na pohyb.

Elekrochemický gradient
 ~ 70 m. V")
Membránový potenciál (klidový) ~ 70 m. V

Nervová buňka - neuron Tělo buňky – umístění jádra a buněčných organel. Dendrity – rozvětvené výběžky, které přijímají signály z axonů. Axon – jediný dlouhý výběžek, který vede signál směrem od těla buňky. Nervový impuls (vzruch) je výsledkem elektrického narušení v plasmatické membráně, které se šíří z jedné části buňky do jiné. Toto elektrické narušení membrány se nazývá akční potenciál a spočívá ve vlně depolarizace membrány pohybující se podél axonu.

Napěťově ovládané Na+ iontové kanály jsou klíčové pro vznik a šíření akčního potenciálu (= vzruchu)

Nadprahová depolarizace způsobí vznik akčního potenciálu. Po obdržení nervového signálu dochází k mírné depolarizaci membrány. Dosáhne-li depolarizace tzv. prahové hodnoty, všechny napěťově ovládané Na+ kanály v dané oblasti se současně otevřou. Na+ vteče dovnitř buňky a způsobí rychlou a velkou depolarizaci membrány. Tato rychlá a velká depolarizace představuje akční potenciál.

Vznik akčního potenciálu vyžaduje depolarizaci membrány nad hodnotu prahového potenciálu. Tento proces začíná na synapsích. presynaptická buňka postsynaptická buňka

Neurotransmitery jsou malé molekuly, které přenáší nervové impulsy na chemických synapsích Figure 21 -28

Jediný neuron kombinuje a vyhodnocuje tisíce excitačních a inhibičních signálů z mnoha dalších neuronů, které ovládají spouštění a frekvenci akčních potenciálů podél axonu. Neurotransmitery uvolněné z každé synapse vyvolají postsynaptický potenciál (PSP). Napěťově ovládané Na+ kanály jsou umístěny na počátku axonu (axon hillock). Akční potenciál vznikne na počátku axonu jen tehdy, když součet všech excitačních a inhibičních PSP v tomto místě přesáhne prahovou hodnotu.
 buňky okolo axonu")
Myelinace zvyšuje rychlost a účinnost šíření akčního potenciálu. Gliální (Schwann cells) buňky okolo axonu vytváří elektrickou izolaci bohatou na glykolipidy. Ranvierovi zářezy rozmístěné v pravidelných intervalech podél axonu jsou místa s přerušenou izolací. Zde se nacházejí napěťově ovládané Na+ kanály. Vtok Na+ v místě jednoho Ranvierova zářezu způsobí depolarizaci v zářezu následujícím v důsledku rychlé difuze Na+ v cytoplasmě. Akční potenciál tímto způsobem přeskakuje z jednoho zářezu na druhý, čímž se značně zvyšuje rychlost a účinnost vedení vzruchu. Tento proces se nazývá saltatorní vedení vzruchu.

Akční potenciál přeskakuje rychle z jednoho zářezu na druhý

The End

Cyklus tří konformací Na+ kanálu – uzavřený, otevřený, inaktivovaný - zajišťuje propagaci akčního potenciálu podél axonu.

Excitační vs. Inhibiční synapse

Čím větší je “kombinovaný PSP”, tím vyšší je frekvence akčních potenciálů vznikajících na počátku axonu (axon hillock). Vlastní hodnota akčního potenciálu je však ve všech případech stejná, protože membránový potenciál je limitován koncentrací Na+ iontů vně buňky.

Neuromuskulární spoj je příklad, jak akční potenciál z neuronu spouští odpověď v jiném typu buňky.

Srovnání přenosu akčního potenciálu přes elektrické a chemické synapse

Vápníková pumpa sarkoplasmatického retikula

Elektrické synapse
- Slides: 69