Sequences Alignment Statistics KaLok Ng Asia University Pairwise





 is the maximum of the")
 =")







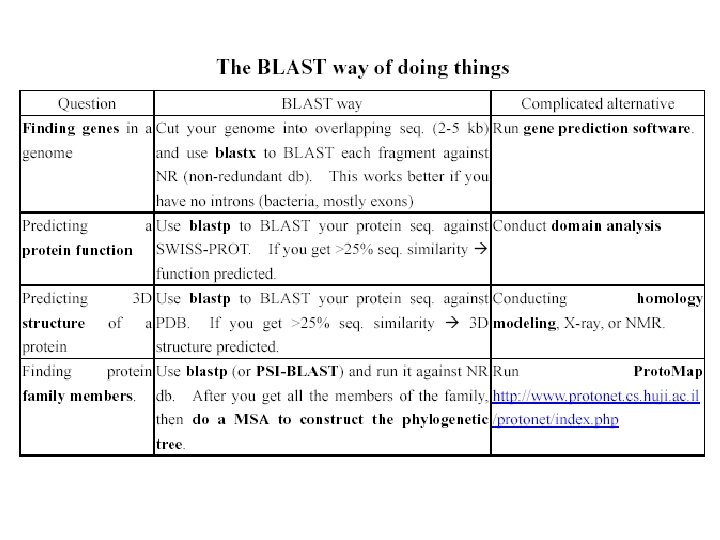
 BLAST - Most highly cited paper >12000 times alternative methods")
 http: //www. ncbi. nlm. nih. gov/BLAST/")
 repeat")

 • How to determine the score scheme ? • Dynamic Programming")
![BLAST (Scoring matrices) Scoring matrices and the Log Odds Ratio where pi[pj] = probability](https://slidetodoc.com/presentation_image/541e70b9f077b6dfb60977fb0a285618/image-20.jpg "BLAST (Scoring matrices) Scoring matrices and the Log Odds Ratio where pi[pj] = probability")
 Amino acid substitution matrix (PAM and BLOSUM) • Leave most adjustable")



 BLOSUM (BLOck SUM) – there are evidence it outperform PAM •")
 The BLOSUM 62 substitution matrix Values below zero indicate amino acid")
 PAM 250 ~ BLOSUM 45 PAM 160 ~ BLOSUM")
")
")
")
")
, Sensitivity (SN), Specificity (SP), and Correlation coefficient (CC) If FP = FN")









, x - ∞, Yev(x) 0, x")

 • BLAST is a fast program quite capable of identifying")
")
 Query sequence – human hemoglobin >gi|57013850|sp|P 69905|HBA_HUMAN Hemoglobin alpha subunit")
 Query sequence – human hemoglobin >gi|57013850|sp|P 69905|HBA_HUMAN Hemoglobin alpha subunit")
 More seqs. are identified than Iteration 1")
 Add or remove the hits that seems to be relevant")
 http: //bioweb. pasteur. fr/seqanal/blast/intro-uk. html")
 B~C")
- Slides: 52
Sequences Alignment Statistics Ka-Lok Ng Asia University
Pairwise Sequence Alignment • The most important class of bioinformatics tools – pairwise alignment of DNA and protein seqs. alignment 1 alignment 2 Seq. 1 ACGCTGA Seq. 2 A - - CTGT ACTGT - Seeks alignments high seq. identity, few mismatchs and gaps Assumption – the observed identity in seqs. to be aligned is the result of either random or of a shared evolutionary origin Identity ≠ similarity Sequence identity/similarity = Homology (a risky assumption) Sequence similarity ≠ Homology
Pairwise Sequence Alignment Same true alignment arise through different evolutionary events Scoring scheme: substitution -1, indel -5, match 3 indel Score 9 5 4 4 Figure A Common evolutionary events and their effects on alignment
Pairwise Sequence Alignment Find the optimal score the best guess for the true alignment Find the optimal pairwise alignment of two seqs. inserted gaps into one or both of them maximize the total alignment score Dynamic programming (DP) – Needleman and Wunsch (1970), Smith and Waterman (1980), this algorithm guarantees that we find all optimal alignments of two seqs. of lengths m and n BLAST is based on DP with improvement on speed
Pairwise Sequence Alignment The score for alignment of i residues of sequence 1 against j residues of sequence 2 is given by where c(i, j) = the score for alignment of residues i and j and takes the value 3 for a match or -1 for a mismatch, c(-, j) = the penalty for aligning a residue with a gap, which takes the value of -5
Pairwise Sequence Alignment • The entry for S(1, 1) is the maximum of the following three events: • S(0, 0) + c(A, A) = 0 + 3 = 3 [c(A, A) = c(1, 1)] • S(0, 1) + c(A, -) = -5 + -5 = -10 [c(A, -) = c(1, -)] • S(1, 0) + c(-, A) = -5 + -5 = -10 [c(- , A) = c(-, 1)] • Similarly, one finds S(2, 1) as the maximum of three values: (-5)-1=-6; 3 -5=-2; and (-10)-5=-15 the best is entry is the addition of the C indel to the A-A match, for a score of -2 (see next page).
Pairwise Sequence Alignment The alignment matrix of sequences 1 and 2 S(2, 1) = max {S(1, 0) + c(2, 1), S(1, 1) + c(2, -), S(2, 0) + c(-, 1)} = max { S(1, 0) + c(C, A), S(1, 1) + c(C, -), S(2, 0) + c(-, A) } = max { -5 -1, 3 -5, -10 -5 } = -2
Pairwise Sequence Alignment Traceback determine the actual alignment From the top right hand corner the (7, 5) cell For example the 1 in the (7, 5) cell could only be reached by the addition of the mismatch A-T ACGCTGA A - - CTGT or ACGCTGA AC - - TGT 4 matches 1 mismatch 2 indels Ambiguity – has to do with which C in seq. 1 aligns with the C in seq. 2
Pairwise Sequence Alignment Parameters settings - Gap penalties • Default settings are the easiest to use but they are not necessarily yield the correct alignment • constant penalty independent of the length of gap, A • proportional penalty is proportional to the length L of the gap, BL (that is what we used in the this lecture) • affine gap penalty gap-opening penalty + gap-extension penalty = A+BL • There is no rule for predicting the penalty that best suits the alignment • Optimal penalties vary from seq. to seq. it is a matter of trial and error • Usually A > B, because of opening a gap (usually A/B ~ 10) • Hint: (1) compare distantly related seqs. high A and very low B often give the best results penalized more on their existence than on their length, (2) compare closely related seqs. , penalize both of extension and extension
BLAST global alignment of a pair of seqs. , in which all residues from both seqs. are included. BLAST – local alignment Interpreting BLAST output -Smith and Waterman algorithm guaranteed to find the best local alignment of two seqs. -Too slow in practice !! -BLAST heuristic search method that is not guaranteed to find the best local alignment, but has been especially effective in practice -e. g. S 45649 (from a fossilized insect) >gi|256517|gb|S 45649. 1| 16 S r. RNA [Mastotermes electrodominicus=termites, amber-preserved fossil, Mitochondrial, 94 nt] AATAAAATTTTAATATAAAGATTTATAGGGTCTTCTCGGCCTTTAAAAA TATTTTAGCCTTTTGAC AAAAAAATCTACAAAAAA
BLAST http: //www. ncbi. nlm. nih. gov/BLAST/ E-value, with the most significant hits listed first E-value is the number of hits with the same level of similarity that you would expect by chance E = 0. 01 occur once every 100 searches even when there is no true match in the database E-value is similar in spirit to the p-value of statistical hypothesis tests. E-value depends on the size of the database. P-value is the probability of finding a seq. similarity as similar as the observed match if there were really no true matches in the database. E-value ≠ p-value E-value ~ p-value when it is small (say < 0. 1) Since we are interested in unusual hits, it is safe to interchange E-value with p-value. E-value – the lower the better the alignment, matches above 0. 001 are often close to the twilight zone Score (bits) – the higher the better the alignment, score below 50 are unreliable
Searching Sequence Databases Using BLAST
Searching Sequence Databases Using BLAST
BLAST The BLAST output may not be the same every time due to the upgrade of several components : Database, the BLAST program, the default parameters of the server E-value, similarity and homology Protein : >25 %, > 100 a. a. , < 10 -4 DNA : >70%, > 100 bp, < 10 -4 Gap penalties - constant penalty independent of the length of gap, A - proportional penalty, penalty is proportional to the length L of the gap, BL - Affine (『數』遠交的, 『化學』親和的) gap penalty, gap-opening penalty + gap-extension penalty = A+BL Remark • Prediction using similarity is a powerful idea in bioinformatics • homologue seqs. evolved by divergence from a common ancestor, therefore to say two seqs. share 50% homology is nonsense; to say two seqs. share 50% similarity and that they indicate possible homology is the correct usage of the terms • Similarity NOT necessary implied homology
BLAST (choosing the parameters) BLAST - Most highly cited paper >12000 times alternative methods seeds + dynamics programming speed up, faster not guaranteed to find the best alignment less accurate
BLAST (Sequence filters) http: //www. ncbi. nlm. nih. gov/BLAST/
BLAST What is a coiled-coil? Coiled-coil domains are characterized by a heptad (成七的一組) repeat pattern in which residues in the first and fourth position are hydrophobic, and residues in the fifth and seventh position are predominantly charged or polar. This pattern can be used by computational methods, such as Multi. Coil (MIT) or SOCKET (University of Sussex)to predict coiled-coil domains in amino acid sequences.
BLAST programs
BLAST (Scoring matrices) • How to determine the score scheme ? • Dynamic Programming do not provide the user with a measure of statistical similarity when regions of local similarity are found • Take into account not just the position-position overlap between two seqs. but the characteristics of the a. a being aligned define scoring matrices • Protein scoring matrices take three major biological factors into account: • Conservation – the numbers within the scoring matrix provide a way of representing what a. a. are capable of substituting for other a. a. (characteristics such as charge, size, hydrophobicity) • Frequency – a. a cannot freely substitute for one another, the matrices need to reflect how often particular a. a occur among the entire proteins. • Evolution – scoring matrices implicitly represent evolutionary patterns, and matrices can be adjusted to favor the detection of closely related or more distantly related proteins.
BLAST (Scoring matrices) Scoring matrices and the Log Odds Ratio where pi[pj] = probability with which a. a i [j] occurs among all proteins qi, j = how often the two a. a i and j are seen to align with one another in MSA of protein families or in seqs. that are known to have a biological relationship.
BLAST (PAM matrices) Amino acid substitution matrix (PAM and BLOSUM) • Leave most adjustable parameters to the default value except the scoring matrix • a simple scheme for scoring seq. matches and mismatches (all mismatches received the same penalty) • Scoring matrix allows some mismatches to be penalized less then others • Leucine-isoleucine mismatch < leucine-tryptophan mismatch • PAM (Point Accepted Mutations) scoring matrices – derived from closely related species (evolutionary point of view, avoid the complications of unobserved multiple substitutions at a single position) • PAM derived from the likelihood of amino acids substitution during the evolutionary process • PAM matrices with a smaller number represent shorter evolutionary distance • PAM 1 – one a. a change per 100 a. a, or roughly 1% divergence
PAM matrix Asp Glu, 0. 95% The values in each column sum to 10000 100%. The value 9730 at the top left indicates that alanine has a 97. 30% chance that the replacement aa will also be an alanine. The least mutable a. a, tryptophan (W) has a 99. 41% chance of remaining the same. The PAM 250 = (PAM 1)250
Construction of a PAM matrix – an example • 1. 2. 3. 4. 5. 6. 7. Construct a MSA. Below is an example of a MSA: ACGCTAFKI GCGCTAFKI ACGCTAFKL GCGCTGFKI GCGCTLFKI ASGCTAFKL GCGCTAFKI ACACTAFKL ACGCTAFKL ASGCTAFKL ACACTAFKL GCGCTGFKI GCGCTLFKI • • • A phylogenetic tree is created on the basis of the alignment. Phylogenetic trees are discussed in any book on bioinformatics. The phylogenetic tree shows various substitutions among the amino acids. For each aa type, calculate its frequency of substitution, and its assume that the substitutions are equally likely in each direction, so a substitution FG, A = A G would also count as a G A substitution. We count all A G and G A branches in the tree. For the tree above, FG, A = 3.
Construction of a PAM matrix – an example • • • Compute the relative mutability, mi, of each aa. For example, consider the A residue. There a total of 4 mutation involving A. Divide this number by the number of mutations in the entire tree time two (6 x 2=12), times the relative freq. of A residue (10 A’s out of 63 residues, i. e. 10/63 = 0. 159), times 100. Thus m. A = 4/12 x 0. 159 x 100 = 5. 3. The value 100 is used so that the PAM-1 matrix will represent 1 substitution per 100 residues in the phylogenetic tree. Compute the mutation probability, Mij , for each pair of aa. is the total number of substitutions involving A in the phylogenetic tree, and it is equals to 4 for A residue. • Finally each Mij is divided by the frequency of occurrence, fi of residue i, and the log of the resulting value becomes the entry Rij in the PAM matrix. For example, f. G = 10/63=0. 1587, i. e. 10 G’s divided by 63 aa. , so RGA = log (3. 975/0. 1587) = 1. 40. • Repeat for each pair of aa PAM matrix
BLAST (BLOSUM matrices) BLOSUM (BLOck SUM) – there are evidence it outperform PAM • Block proteins in the same family can be aligned without introducing a gap (not the individual seqs. ) • So any given protein can contain one or more blocks, corresponding to each of its functional or structural motif • With these protein blocks, it is possible to look for substitution patterns only in the most conserved regions of a protein block substitution matrices are generated • BLOSUM scoring matrix – based on data from distantly related seqs. (default BLOSUM 62 for general use) • The most commonly used matrices are PAM 120, PAM 250, BLOSUM 50 and BLOSUM 62 • BLOSUM matrices with a smaller number represent a longer evolutionary distance
BLAST (BLOSUM matrices) The BLOSUM 62 substitution matrix Values below zero indicate amino acid changes that are more likely to have a functional effect than values of zero and above.
BLAST (relating PAM to BLOSUM) PAM 250 ~ BLOSUM 45 PAM 160 ~ BLOSUM 62 PAM 120 ~ BLOSUM 80 Selecting an appropriate scoring matrix Matrix Best use Similarity(% ) BLOSUM 90 Short alignments that are highly similar 70 -90 BLOSUM 80 Detecting members of a protein family 50 -60 BLOSUM 62 Most effective in finding all potential similarities 30 -40 BLOSUM 30 Longer alignment of more divergent seqs. <30
BLAST (Sensitivity and Specificity)
BLAST (Sensitivity and Specificity)
BLAST (Sensitivity and Specificity)
BLAST (Sensitivity and Specificity)
Accuracy (Q), Sensitivity (SN), Specificity (SP), and Correlation coefficient (CC) If FP = FN = 0, CC =1 (the prefect prediction), If TP = TN = 0, CC = -1 (the worst prediction)
BLASTing DNA sequences
Use of BLASTx to find ORF AE 008569
Use of BLASTx to find ORF Frame = +1 Frame = -2
Use of BLASTx to find ORF
Use of BLASTx to find ORF
Use of BLASTx to find ORF
The BLAST algorithm http: //cbsu. tc. cornell. edu/resources/seq_comp/webtutorials/ Outside_tutorials. html
The BLAST report
The Gumbel Extreme Value Distribution • • • Test the significant of a local alignment score For simplicity, consider un-gap alignment Base on the distribution of scores expected by aligning two random seqs. of the same length and base composition (nucleotides or a. a) as the two test seqs. • These random seq. alignment scores follow a distribution called the extreme value distribution • Somewhat like a normal distribution with a positively skewed tail in the higher score range Goal • To valuate the probability that a score between random or unrelated seqs. will reach the score found between two real seqs. of interest. If that probability is very low, the alignment score between the real seqs. is significant. • Reference: Mount D. W. Bioinformtics, CSHL, 2001.
The Gumbel Extreme Value Distribution Behavior of Yev(x), x - ∞, Yev(x) 0, x ∞, Yev(x) 0 x =0, Yev(x) e-1 The expectation value or mean of x is the value of the Euler-Massceroni constant, g = 0. 57722. . , and the variance of x, s 2 is p 2/6 = 1. 6449. The probability that score S less than value x, P(S<x) is obtained by calculating the area under curve from –∞ to x,
The Gumbel Extreme Value Distribution To facilitate calculation, a sequence alignment score S is normalized to produce a score S’. Example Length of query : 250 Length of random query : 250 For PAM 250 matrix, Lambda : 0. 229, K : 0. 09 The raw score: 75 : bit-score S' = 0. 229*75 – ln(0. 09*250) S’ = 8. 54 bits P(S’ ≧ 8. 54) = e-8. 54 = 1. 99*10 -4 This means that the chance that an alignment between two random sequences will achieve a score greater than or equal to 75 using the PAM 250 matrix is 0. 000199. E-value = P*size of database
Position-Specific Iterated BLAST (PSI-BLAST) • BLAST is a fast program quite capable of identifying the close matches, but it is less sensitive for remote homologous seq. search • For example, find the homologous of a mouse protein in the human genome using BLAST is easy • But if you want to search for a homologous of a yeast protein in the human genome (remote homologous seq. ), the job is more difficult for BLAST
Position-Specific Iterated BLAST (PSI-BLAST)
Position-Specific Iterated BLAST (PSI-BLAST) Query sequence – human hemoglobin >gi|57013850|sp|P 69905|HBA_HUMAN Hemoglobin alpha subunit (Hemoglobin alpha chain) (Alpha-globin) MVLSPADKTNVKAAWGKVGAHAGEYGAEALERMFLSFPTTKTYFPHFDLSHGSAQVKGHGKKVADALTNAVAHVDDMPN ALSALSDLHAHKLRVDPVNFKLLSHCLLVTLAAHLPAEFTPAVHASLDKFLASVSTVLTSK YR 0 ≦E-value < 10 -40
Position-Specific Iterated BLAST (PSI-BLAST) Query sequence – human hemoglobin >gi|57013850|sp|P 69905|HBA_HUMAN Hemoglobin alpha subunit (Hemoglobin alpha chain) (Alpha-globin) MVLSPADKTNVKAAWGKVGAHAGEYGAEALERMFLSFPTTKTYFPHFDLSHGSAQVKGHGKKVADALTNAVAHVDDMPNALSALSDLHA HKLRVDPVNFKLLSHCLLVTLAAHLPAEFTPAVHASLDKFLASVSTVLTSK YR Gene or Structure information
Position-Specific Iterated BLAST (PSI-BLAST) More seqs. are identified than Iteration 1
Position-Specific Iterated BLAST (PSI-BLAST) Add or remove the hits that seems to be relevant or irrelevant (non-human seq. )
Position-Specific Iterated BLAST (PSI-BLAST) http: //bioweb. pasteur. fr/seqanal/blast/intro-uk. html
Position-Specific Iterated BLAST (PSI-BLAST) B~C