Sejtmembrn szerkezete s mkdse Dr habil Khidai Lszl

Sejtmembrán szerkezete és műküdése Dr. habil. Kőhidai László egyetemi docens SE, Genetikai, Sejt- és Immunbiológiai Intézet 2008.

Prokaryota sejt citoplazma Eukaryota sejt tok szekréciós szemcse sejtfal sejtmembrán csillók mikrobolyhok pinocitotikus vezikulum riboszóma sejtmembrán mikrotubulus thylakoid pílusok ostor mikrofilamentum színtest centriolumok Golgi apparátus sima felszínű endoplazmatikus hálózat riboszóma lizoszóma sejtmagvacska kromatin állomány mitochondrium nukleoplazma citoplazma durva felszínű endoplazmatikus hálózat pórusok sejtmag- sejtmag membrán

Prokaryota sejt citoplazma Eukaryota sejt tok szekréciós szemcse sejtfal sejtmembrán csillók mikrobolyhok pinocitotikus vezikulum riboszóma sejtmembrán mikrotubulus thylakoid pílusok ostor mikrofilamentum színtest centriolumok Golgi apparátus sima felszínű endoplazmatikus hálózat riboszóma lizoszóma sejtmagvacska kromatin állomány mitochondrium nukleoplazma citoplazma durva felszínű endoplazmatikus hálózat pórusok sejtmag- sejtmag membrán

Kompartmentalizálódás Az egyes biokémiai folyamatok membránokkal határolt terekbe történő szétválasztása – optimális környezeti feltételek biztosítása

A citoplazmatikus membránok fő funkciói Határhártya képzése – tápanyagok be és kifelé irányuló tarnszportját segíti elő Fehérjék kihorgonyzása – a traszport, bioenergetikai folyamatok, szignálok észlelése és kemotaxis Energia raktározás - proton gradiens generálása
 –")
A membránok funkciói • 1. Határhártyát képeznek és szelektív átjárhatóságú felszínek (ld. permeabilitás) – sejt membrán/plazmamembrán és intracellularis membránok – sejt organellumok • 2. Specifikus funkciókat töltenek be – a kapcsolódó proteinek alapján megkülönböztethetők (integráns vagy perifériás fehérjék). Glucose phosphatase – az ER-membrán-kötött enzym és egyben annak markere is. • 3. A membránok szabályozzák egyes anyagok transzportját – a. Egyszerű diffúzió (magas cc. ->alacsony cc. ): H 2 O, O 2, CO 2, ethanol b. Facilitált diffúzió (magas cc. ->alacsony cc. ) transzport proteinek segítségével: cukrok, aminosavak c. “Pumpa mechanizmusok” proteinjei, melyek az ion-gradienseket tartják fenn: alacsony cc. -> magas cc. Cukrok, aminosavak sejtbe juttatása koncentráció gradiens ellenében is. d. Endocytosis és exocytosis, stb

A membránok funkciói • 4. Elektromos és kémiai jelek detektálása és továbbítása – szignál transzdukció: külső jelek fogadása és továbbítása. Pl. : Ösztrogén – hidrofób karaktere miatt képes átjutni a membránon; más anyagok hatásához receptorhoz való kötődés szükséges, mely másodlagos hírvivőket generál. A jel a sejtmagi szintig is eljuthat és befolyásolhatja a génexpressziót. • 5. Sejt-sejt kommunikáció közvetítése – gap junction (állati sejtek) és plasmodesmata (növényi sejtek) intercelluláris kommunikációt tesznek lehetővé.

Membrán-modellek Lipid-oldékony anyagok gyorsan bejutnak a sejtbe Benzin-lipid keverék, a benzin elpárolgása után molekuláris lipid film keletkezik Benzin-oldékony lipidek két rétegből álló filmet alkotnak víz felszínén A fehérjék integráns részei a sejt membránnak. A lipid bilayert kívül-belül proteinek rétege fedi. Részben magyarázza a proteinek, cukrok, ionok és más hidrofil anyagok gyors átjutását. Elektronmikroszkóp felfedezése. A sejtek plazmamembránnal burkoltak. „Unit-membrane”modell. Proteinek mozaikos elrendeződése a membránban. Egyes proteinek átérik a membránt, transzmembrán proteinek

Fluid mozaik membrán modell Singer – Nicolson 1972 vvt. membrán

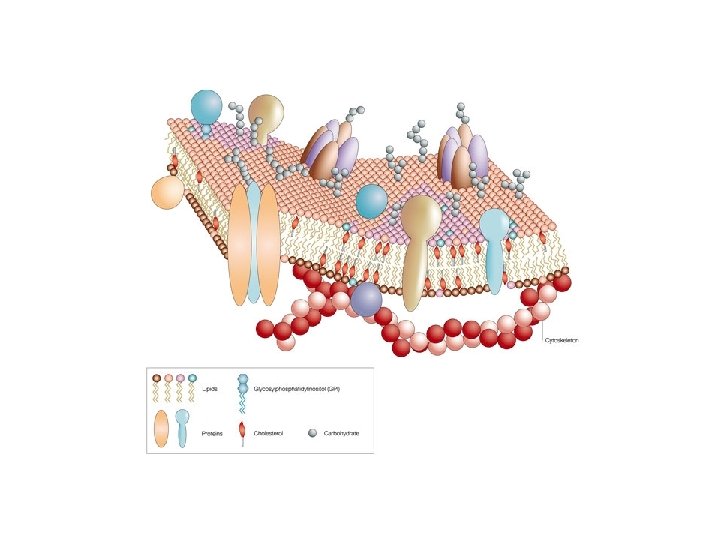
Membrán modellek – Folyékony mozaik modell: A proteinek mozaikosan, elszórtan és beágyazottan helyezkednek el a folyékony lipid kettősrétegben Bacteriorhodopsin: az első transzmembrán doménnel leírt protein, mely retinol-szerű részével energia megkötő képességű is egyben. A napfény energiája megkötődés után szerkezeti változást idéz elő, mely protonok sejtből tőrténő kijuttatását eredményezi. A kialakuló protongradiens energia-forrásként szolgál. A legtöbb membrán felépítésében lipidek, fehérjék és szénhidrátok is részt vesznek.

Foszfolipidek általános szerkezeti felépítése
 Cholesterin Cerebrozid Szfingolipidek")
A membránt felépítő fő lipid-komponensek Foszfolipidek Glikolipidek Egyéb membrán-lipidek (nem foszfolipidek) Cholesterin Cerebrozid Szfingolipidek Ceramid

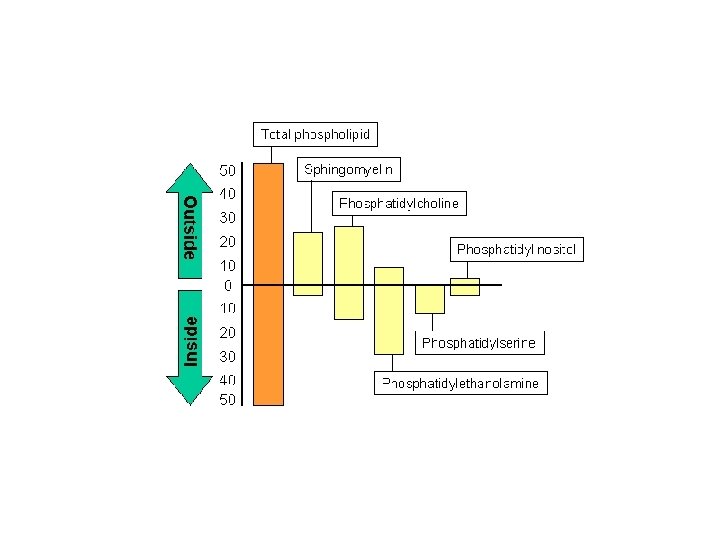
Membrán lipidek: a modell „fluiditásáért” felelős elemek A membránok alkotásában több féle lipid vesz részt • Foszfolipidek Foszfogliceridek Szfingolipidek Különböző membránok foszfolipid összetétele

• Glikolipidek Cerebrozidok és gangliozidok Tay-Sachs betegség: β–N-acetilhexoaminidáz hiányos lizoszómák, gangliozidok degradációja károsodik; gangliozid akkumuláció az idegrendszerben. • Szteroidok (nem található a prokaryotákban vagy a mitochondrium, színtest belső membránjaiban) Cholesterin Fitoszterin A baktériumok és a cyanobaktériumok membránjaiban szteroid-szerű molekulák találhatók – haponoid (petroleum depozitokban gazdag; korai prokaryoták membrán componensei? )

Vékonyréteg-kromatográfia – a lipid analízis fontos eszköze: Álló fázis – vékony szilikagél, hidrofil rétege üveg vagy fém lemezen Mozgó fázis – oldószer chloroform, methanol, és water keveréke Alacsony affinitás Cholesterin Phosphatidylethonolamine (PE) Phosphatidylcholine (PC) Phosphatidylserine (PS) Magas affinitás Zsírsavak a membrán szerkezete és működése szempontjából is jelentősek A zsírsavak minden membrán-lipid alkotói kivéve a szteroidokat. A hidrofób szénhidrogének általában 16 - vagy 18 -C hosszúak és a lipid kettősréteg stabilitását adják. Telített zsírsavak: palmitinsav(16 C) és sztearinsav (18 C) Telítetlen zsírsavak: Olajsav és linolsav (mk. 18 C)

Membrán- aszimmetria: a lipidek döntő hányada egyenlőtlen arányban oszlik meg a két lipid-monolayerben - mennyiségi és minőségi eltérések Az aszimmetria a lipidek típusainak eltérőségében és a telítettség fokának különbözőségében testesül meg. Az aszimmetria a membránok biogenezise során alakul ki. A kialakult aszimmetria termodinamikai okoból gátolja a lipidek cseréjét a két felszín között. Külső membrán: glikolipidek – felismerő folyamatok és jeltovábbítás Belső membrán: a sejt belső terébe történő jeltovábbításban fontosak -foszfatidiletanolamin, foszfatidilinozitol, foszfatidilszerin Lipidek mozgása a membránban: • “flip-flop”, vagy transzverz diffúzió • rotáció • laterális diffúzió A flip-flop mozgás ritka, de a biológiai membránokban az ú. n. foszfolipid transzlokátorok vagy flippázok segítik elő. Ezért gyakoribb a foszfolipid transzlokáció a biológiai membránokban, mint a mesterséges lipid bilayerekben.

A membrán aszimmetria kialakulása és fenntartása: § Foszfolipidek szintézise az ER membránhoz kötötten zajlik. § A foszfolipidek az ER citoplazmatikus felszínéhez kapcsolódnak § A flippáz juttatja át az egyes lipideket a bilayer másik oldalára.

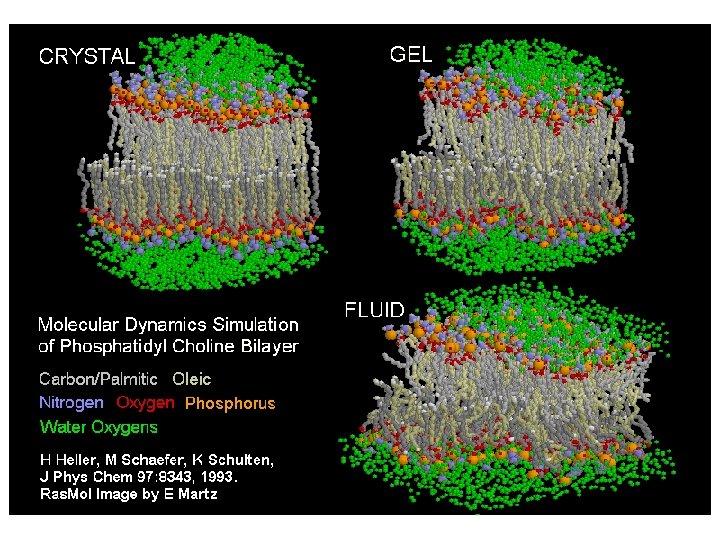
A lipid bilayer fluiditásának vizsgálata Laterális diffúzióval a lipidek és proteinek pl. : egy 800 M. S. -jú lipid néhány m. M (egy baktérium hosszával azonos távolságot) tud elmozdulni 1 mp alatt! A proteinek nagy méretük miatt lassabban mozognak. Vizsgálati eljárás: fluorescence recovery after photobleaching (FRAP) technika A membrán megfelelően csak „fluid” állapotában működik Fázis-átmeneti hőmérséklet (Tm): Fázis-átmenet: gél-szerű állapot -> fluid „Differential scanning calorimetry” Calometer: a hőabszorpció mérésére szolgáló kamra. A görbe csúcsának helye jelzi a Tm-et. Telítetlen: alacsony Tm Telített: magas Tm

76 o. C Membrán-fluiditás és zsírsavösszetétel kapcsolata Két fő szempont: lánchossz és telítettség mértéke Membrán-lipidek gyakran 1 telített és 1 telítettlen zsírsavból épülnek fel 70 o. C 32 o. C Szteroidok hatása a membránfluiditásra -11 o. C Szteroidok – cholesterin és phytosterin. A szteroidok a teljes membrán-lipid tartalom 50%-át is kitehetik Cholesterin – hidrogén kötéseket alakít ki a szomszéd foszfolipidek oxigén atomjaival. A cholesterin paradox hatása: magas hőmérsékleten csökkenti a membrán fluiditást (a rigid gyűrűk hatása) míg alacsony hőmérsékleten növeli a fluiditást (a szomszédos szénhidrogén láncok természetes elrendeződését gátolva) AZONBAN, AZONBAN a szteroidok csökkentik a lipid bilayer permeabilitását – a foszfolipidek közötti teret kitöltve. Az élőlények szabályozni képesek a membrán fluiditást Poikelotherm-ek (baktérium, gomba, növény, kétéltűek stb. ) vs. homeotherm-ek (melegvérű állatok) Homeoviszkózus adaptáció – a membrán viszkozitása megtartott a változó hőmérséklet ellenére – terminális C-k eltávolításával zsírsavláncok rövidítése, ill. telítetlenné tétele (pl. : E. coli desaturáz). 18 -C lipidek

A membrán fluiditásának befolyásolása: Foszfolipidek: lánchossz telítettség Cholesterin-tartalom

Lipid raft-ok: - cholesterinben gazdag - mikro-domainek -glikolipidek+szfingolipidek - a raftok jelen vannak a sejt felszíni membránján és a Golgiról lefűződő membránokon - jelentőségük: sejtmembrán egyes rétegei közötti szabadenergia különbség csökkentése - membránhoz kötött transzport folyamatok Más nevei a szakirodalomban: -detergent-insoluble glycolipid-enriched complexes (GEM vagy DIG) - detergent resistant membranes (DRMs). Kimutatás: fluoreszcens mikroszkópia – GM 1 gangliozidok; AFM
 után gyémánt-késsel történő feltárás")
Membrán-proteinek: a modell “mozaik”-jellege Freezing fracturing: gyors fagyasztás (folyékony nitrogén) után gyémánt-késsel történő feltárás E felszín (exoplazmatikus oldal) és P felszín (protoplazmatikus oldal)

Membrán proteinjei: Integráns membrán proteinek: ~20 aminosavból álló, transzmembrán szegmentum. Nehezen izolálható detergens kezelés kell az extrakciójához. . Intergráns monotóp proteinek Transzmembrán proteinek: singlepass multi-subunit - integráns - perifériális - lipidek által kihorgonyzott Glycophorin (vvt. ) Bacteriorhodopsin Hydropathy plot

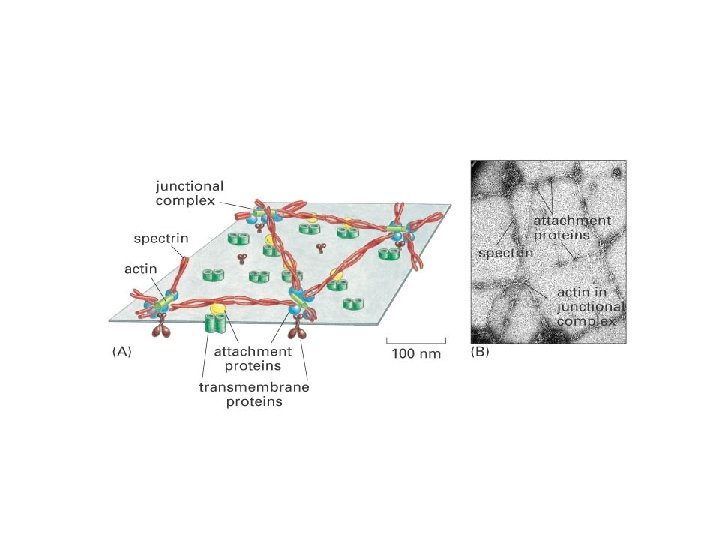
Perifériásl membrán proteinek a membrán felszínekhez gyenge elektrostatikus erők vagy hidrogén kötések rögzítik. Kapcsolat az integráns proteinek hidrofil részeivel vagy a membrán lipidek poláris feji részeivel. pl. : spectrin és ankyrin a vvt. plazmamembránjában. A proteineket SDS-polyacrylamid gél elektroforézissel választhatjuk el Membrán proteinek izolálása Perifériás membrán proteinek: a p. H vagy az ionos kötések megváltoztatása, calcium eltávolítása –chelatiós-agensek (kationos-kötés), vagy urea (a hidrogén kötéseket bontja). A lipid-kötött proteinek hasonló módon izolálhatók, mint a perifériás proteinek, mivel itt is kovalens kötéseket kell bontani. Az integráns membránproteinek kivonása nehezebb, detergens igényes feladat. A detergens a hidrofób kötéseket bontja és a lipid bilayerrel lép kölcsönhatásba ld. SDS (sodium dodecyl sulfate) kezelés SDS-polyacrylamid gél elektroforézis: Elektroforézis elektromos tér alkalmazása elektromosan töltött molekulák elválasztására. A vándorlási sebesség a töltéstől és a mérettől függ. SDS feltárás (proteinek a negatívan töltött SDS-sel fedettek) -> elektroforézis -> festés Coomassie brilliant blue-val. Lipidekhez kötött membrán proteinek: kovalens kötés a lipiddel. • Zsírsav vagy prenyl mint kihorgonyzó • GPI-kapcsolófdás (glycosylphosphatidyinositol, GPI)

Membrán-proteinek izolálása és vizsgálata: Az alkalmazott detergensek a proteinek körüli foszfolipid szerkezetet utánozzák hidrifób hidrofil
")
A membrán-proteinek számos funkciót tölthetnek be Enzim: Pl. : glukóz foszfatáz (az ER-mal asszociált) és glicerinaldehid-3 - foszfát dehidrogenáz (GPD), a vvt. Perifériás proteinjei a vér glukóz-metabolizmusában vesznek részt. Transzport proteinek pl. : transzport ATPázok ATP energiáját felhasználva juttatnak át ionokat a membránon. Receptorok: pl. : hormonok és növekedési faktorok Egyéb: endocitóziss és exocitózis; targeting, szortírozás, fehérjék módosítása az ER vagy a Golgi-ban; fény érzékelése; a sejtmembrán szerkezetének stabilizálása és alak meghatározás (ankyrin, spectrin). TRANSZPORTER KIHORGONYZÁS Transzmembrán (intergráns) Single-pass Multi-pass ? ? ? RECEPTOROK ENZYMEK Membrán-asszociált (perifériás) Kovalensen kötött vs nem-kovalens

A membrán-proteinek bilayerbe ágyazódása Az a-helikális szerkezet igen gyakori. Ennek oka, hogy az így expresszált hidrofób aminosavak felszínre kerülésével a proteinek jól rögzülnek a membrán hidrofób rétegében Az a-helikális szerkezetek hidrofil komponensei a pórusképzésben fontosak

A sejtmembrán szénhidrát komponensei Funkciók: • Felszíni védelem • Felismerés • Sejtadhézió -- extracellularis matrix

A membrán-proteinek nagy része glikozilált Emberi vvt. membrán: 52% proteinek, 40% lipidek és 8% szénhidrátok. A szénhidrátok glikolipid (ritka) és glikoprotein (gyakori) formában találhatók a membránban Glikoproteinek: N-kötöttek az Asp oldalláncának amino csoportján keresztül O-kötöttek – Ser vagy Thr hidroxil csoportjaihoz kötött szénhidrátok Egyes O-kötött szénhidrát hidroxi-Lys vagy hidroxi-Pro hidroxil csoportján keresztül kapcsolódik

A proteinekhez kapcsolódó szénhidrátok hossza: 2 -60 egység, egység egyenes v. lineáris Gyakori felépítő elemek: galaktóz, mannóz, N-acetilglukózamin és sziálsav A vvt. integráns proteinjének (glycophorin) szénhidrát láncainak (16 ilyen láncból 15 Okötött és 1 N-kötött) terminális pozíciójában negatívan töltöt sziálsav található – ez csökkenti a vvt. -k összetapadási hajlamát és a vér viszkozitását. A glikoproteinek elsődleges szerepet töltenek be a sejtmembránok sejt-sejt felismerő funkciójában. Kísérletek: Lectinek: növényi (állati) fehérjék, melyek cukorspecifikus kötést alkotnak. Pl. : wheat germ agglytinin (búza) N-acetilglukózaminra végződő oligoszaccharidokhoz kötődik, míg a „jack bean”-ből kivont concanavalin az oligoszaccharidok belső mannóz csoportjait ismeri fel. Ferritin, egy vas-tartalmú fehérje, melyet glycoproteinekhez kötnek és elektrondenzitásuk révén a membránhoz való kötődés így jól detektálható.

A membránproteinek mobilitása igen eltérő lehet Egyes proteinek szabadon elmozdulnak, míg mások kötöttek. A mobilitás kimutatására felhasznált technika: sejtfúzió Membrán proteinek Egér sejt Emberi sejt Membrán proteinek jelölése fluoreszcensen jelzett antitesttel Vírus indukálta hibrid sejt fúzió A proteinek néhány perc múlva keveredni kezdenek 40 perc múlva a keveredés tejles

A sejtmembrán proteinjeinek mozgása korlátozott: Pl. bélhámsejtek vagy mirigyhám polarizált felszíne Receptorok, transzporterek, sejtkapcsoló struktúrák polarizált elhelyezkedése, megjelenése.

A membránok dinamikus jellege bilayer 5 nm • Számos sejtélettani folyamat során változik a membrán alakja • Az alkotó elemkre ható erők módosítják a membrán kiterjedését. • A komponensekre ható feszítő és nyújtó-erők magyarázzák a mebrán alakváltozásait. • A membránhoz kötött enzimek fenti hatásokat módosíthatják.

A membrán nyújtása A membrán hajlítása RELAXÁLT ÁLLAPOT konstans terület/fej NINCS nyújtás NEM-RELAXÁLT ÁLLAPOT Konstans lipid denzitás Nyújtás - feszülés Lipid denzitás d Membrán aszimmetria

A diffúzió és a hajlat kialakulás egyaránt az elasztikus stressz csökkentését eredményezi Flippáz (Aminophospholipid translocase) Kontrakció Nyújtás

Konzerválás A perturbáció foka Átfordulások száma A denzitás és a hajlatok egységnyiek a deformált membránra nézve Oldószer-áram Membrán-áram

A membránba beépülő elemek pl. proteinek “spontán hajlatok”-at idéznek elő HA A RIGIDITÁS <0 A MEMBRÁN SPONTÁN HAJLATOKAT KÉPEZ

Összefoglalás

Hőmérséklet Szilárd Folyékony

Extracelluláris Intracelluláris

Források: Hardin, J. , Kleinsmith, L. , Becker, W. : World of the cell with free solutions, (Benjamin Cummings, 2002) Alberts, B. , Johnson, A. , Lew, J. : Essential Cell Biology, (Garland, 2004)
- Slides: 45