Regolazione del catabolismo dei carboidrati Regolazione delle vie

Regolazione del catabolismo dei carboidrati

Regolazione delle vie metaboliche La regolazione avviene attraverso: interazioni allosteriche con effettori allosterici, intervento di ormoni, biosintesi di enzimi Molti enzimi di regolazione sono situati in punti critici di ramificazione del metabolismo Regolazione via catabolica e corrispondente via anabolica Glicolisi-gluconeogenesi Gli enzimi regolativi sono regolati in modo reciproco da effettori allosterici comuni Si evitano i cosiddetti cicli futili
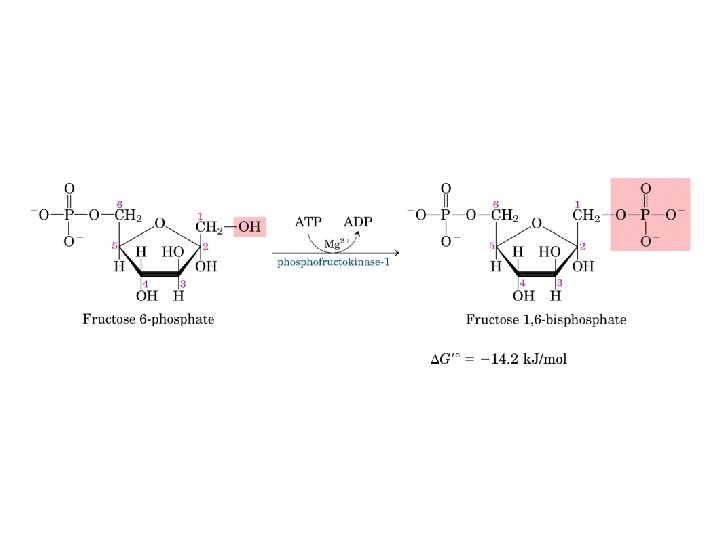
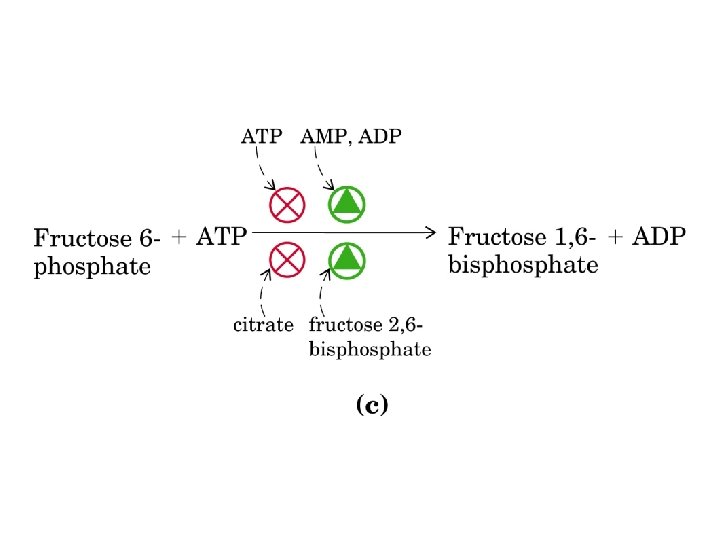

Fruttosio 2, 6 -bisfosfato è un attivatore della via glicolitica ed un inibitore della via gluconeogenetica

Nel 1980 il fruttosio 2 -6 bisfosfato fu identificato come un potente attivatore della fosfofruttochinasi I, sposta l’equilibrio T-R verso R PRODOTTO DALL’ENZIMA: FOSFOFRUTTOCHINASI 2

Enzima bifunzionale

Il glucacone secreto dal pancreas segnala la carenza di glucosio nel sangue

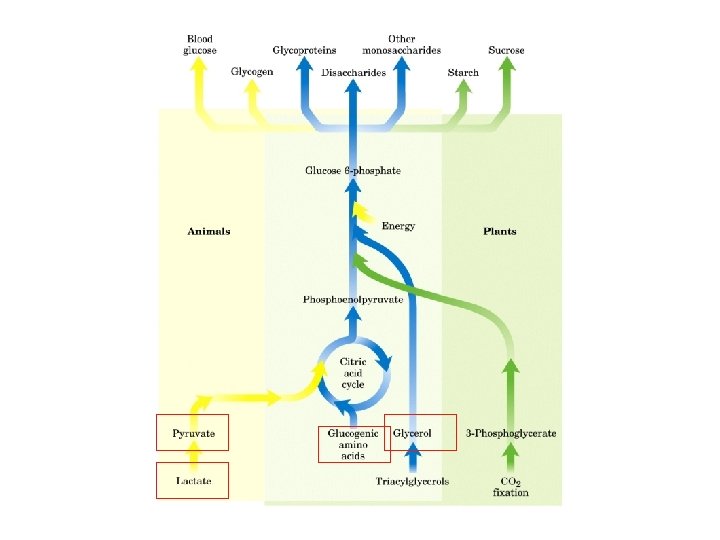
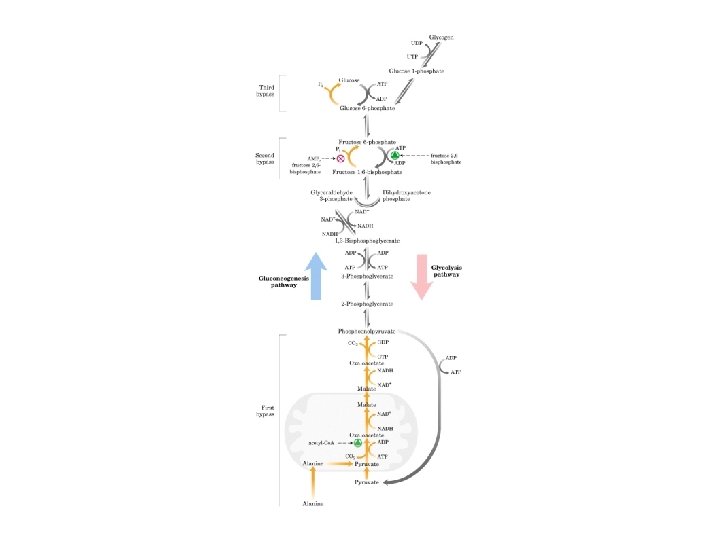
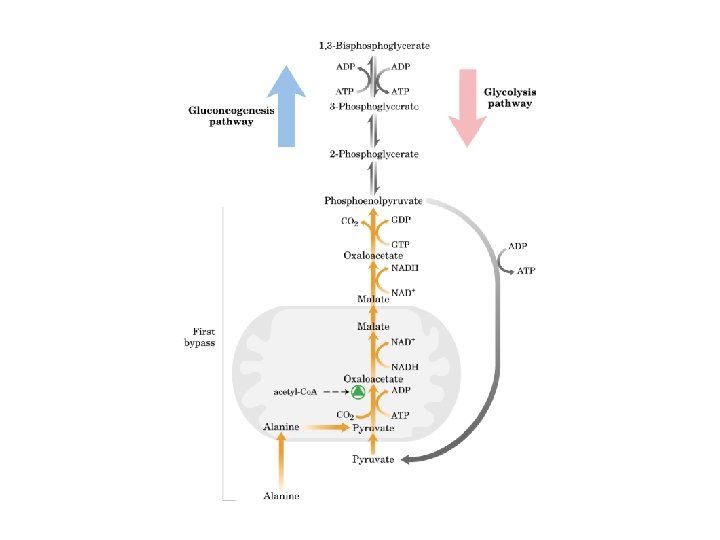
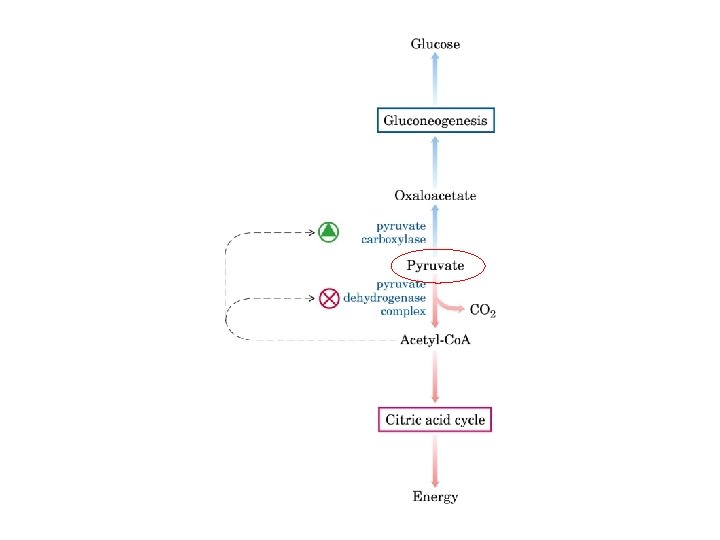
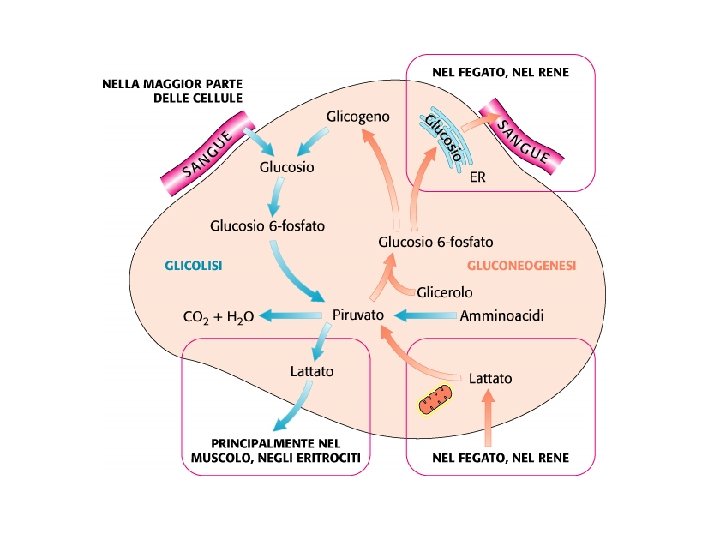
Gluconeogenesi sintesi di glucosio a partire da precursori non glucidici avviene soprattutto nel fegato in misura minore nell’encefalo, muscolo scheletrico e muscolo cardiaco La principale funzione è di mantenere i livelli di glucosio ematico sufficientemente alti da consentire all’encefalo e al tessuto muscolare di avere sufficienti quantità di glucosio Tale processo avvine in tutti gli animali, piante, funghi e microorganismi. Le reazioni sono sostanzialmente le stesse in tutte le specie. Negli animali, i precursori del glucosio sono i composti a tre atomi di carbonio come il lattato, il piruvato, e il glicerolo, oltre ad alcuni amminoacidi. La gluconeogenesi e la glicolisi non sono vie metaboliche identiche percorse in direzioni opposte, anche se condividono diverse tappe.

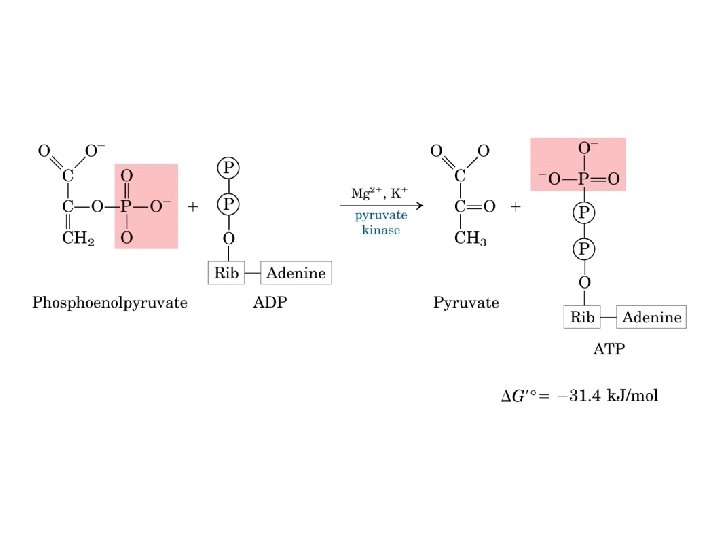
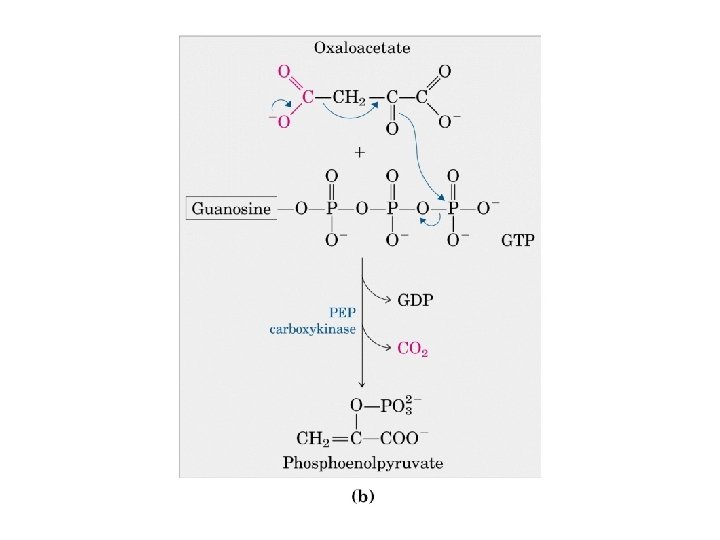
Gluconeogenesi e glicolisi • 7 delle 10 reazioni enzimatiche della gluconeogenesi sono reazioni della glicolisi che avvengono nella direzione opposta • 3 reazioni della glicolisi sono irreversibili e non sono utilizzabili nella gluconeogenesi: - Conversione del glucosio in glucosio 6 fosfato - Fosforilazione del fruttosio 6 fosfato a fruttosio 1, 6 bifosfato - Conversione del fosfoenolpiruvato in piruvato - Nella gluconeogensi queste 3 tappe sono superate mediante un diverso gruppo di enzimi
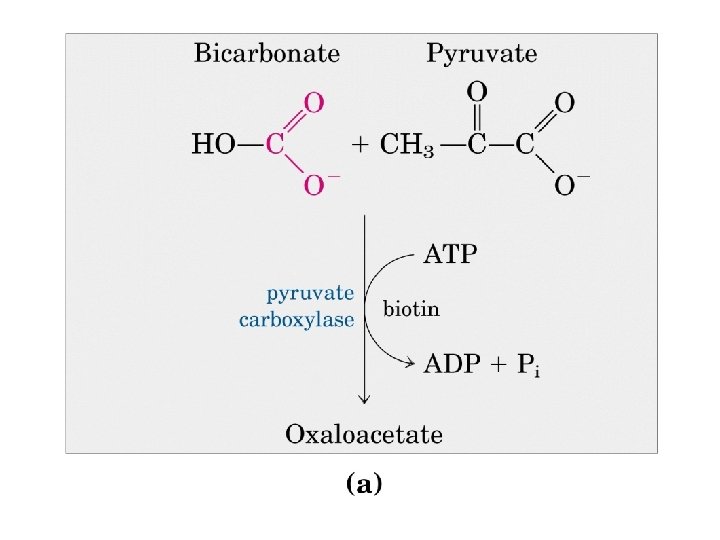

Biotina: trasportatore di gruppi ad un atomo di carbonio nella forma più ossidata Carenze di biotina in soggetti che mangiano grandi quantità di uova crude

Strutturalmente la biotina presenta due anelli tra loro condensati: uno tiofenico ed uno imidazolico. Legata all'anello tiofenico, vi è una catena laterale di acido valerianico

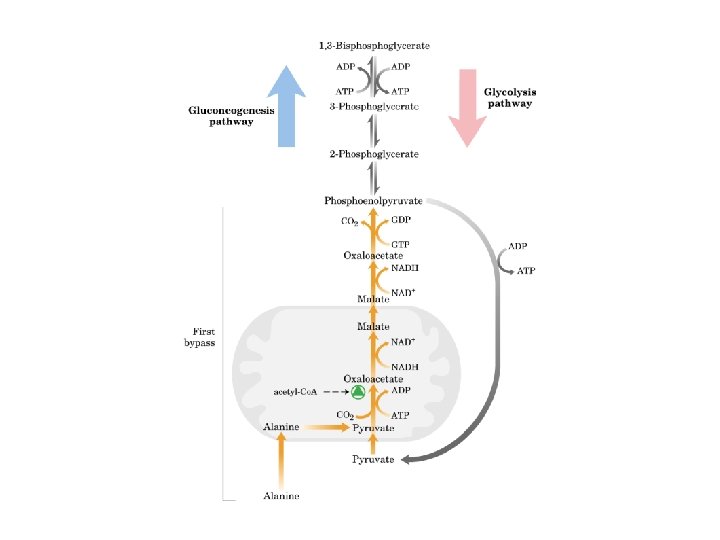
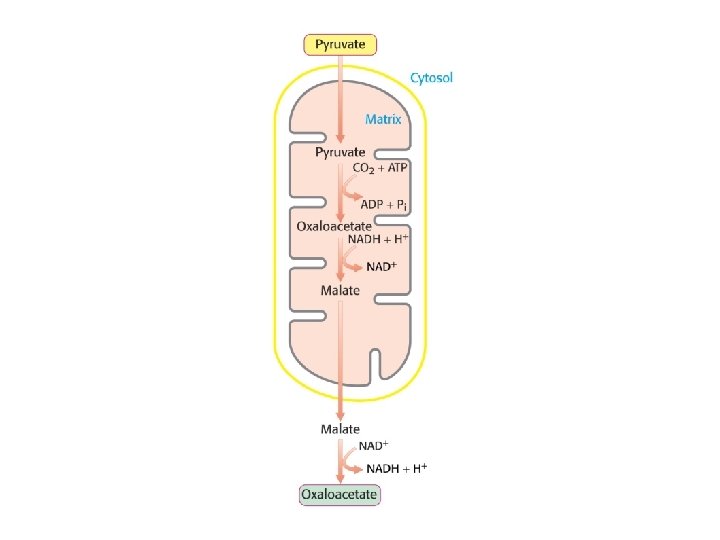
La piruvato carbossilasi è mitocondriale mentre gli altri enzimi della via gluconeogenetica sono citoplasmatici

PERCHE’ LA GLUCONEOGENESI HA BISOGNO DI UN PASSAGGIO ATTRAVERSO I MITOCONDRI? IL RAPPORTO NADH/NAD+ NEL CITOSOL E’PIU’ BASSO DI QUELLO DEI MITOCONDRI NEL CITOSOL IL NADH TENDE A SCARSEGGIARE
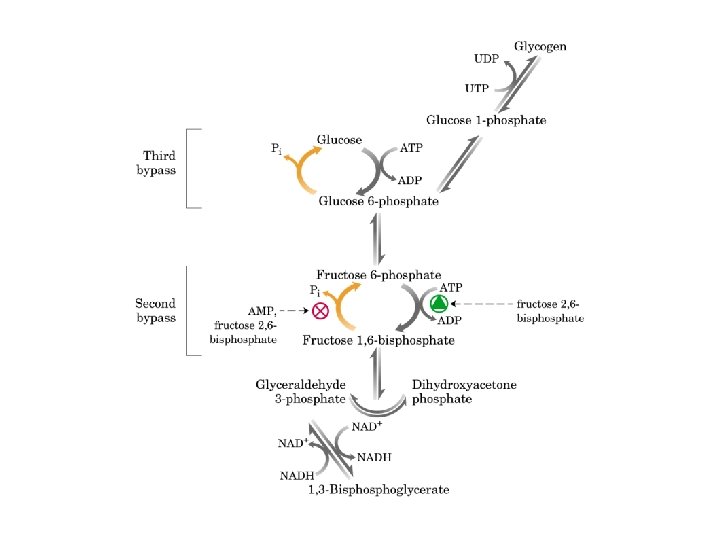

GLUCOSIO-6 -FOSFATASI ASSENTE NEL MUSCOLO E NEL CERVELLO FRUTTOSIO 1. 6 BISFOSFATASI

2 PIRUVATO + 4 ATP + 2 GTP +2 NADH + 4 H 2 O GLUCOSIO + 4 ADP + 2 GDP + 6 P I + 2 NAD+ + 2 H+i

LA GLUCONEOGENESI RICHIEDE L’IDROLISI DI 4 ATP e 2 GTP La glicolisi di una molecola di glucosio produce 2 ATP Per cui per sintetizzare glucosio da una molecola di piruvato bisogna consumare 4 molecole ad alto potenziale di trasferimento del gruppo fosforico

Nella maggior parte dei tessuti la gluconeogenesi termina a glucosio 6 -fosfato Solo nel fegato, che ha il compito di mantenere l’omeostasi del glucosio ematico, il glucosio 6 -fosfato è convertito in glucosio
- Slides: 30