PSY 2055 Psychologie de la perception Vision spatiale

 droit lumière 1 - Corps genouillés latéraux")
 • Corps genouillé latéral : Noyau du thalamus. Lieu de")


 • Les champs récepteurs des neurones du")
 Chaque LGN a 6 couches (la")
 Chaque couche d’un LGN est rétinotopique. Les neurones situés")
 • Les couches 1 et 2 sont")
 • Les cellules des couches magnocellulaires")
 droit lumière Corps genouillé")
")
")







 varie en")









 • Lorsqu’une électrode suit une trajectoire oblique, les champs récepteurs des")






 provoque un effet")










! = + réseau Analyse de Fourier 2 D :")
! = + Fréquence = 1 + + Fréquence =")
!")
")




- Slides: 59
PSY 2055. Psychologie de la perception. Vision spatiale : d’étoiles à grilles Frédéric Gosselin
Plan du cours Corps genouillé latéral (LGN) droit lumière 1 - Corps genouillés latéraux Corps genouillé latéral (LGN) gauche 2 - Cortex visuel
Corps genouillés latéraux (CGL) • Corps genouillé latéral : Noyau du thalamus. Lieu de synapse reliant le nerf optique et le cortex visuel. – Le CGL gauche reçoit de l’information provenant du côté gauche de la rétine et ce, pour les deux yeux (champ visuel droit). – Le CGL droit reçoit de l’information provenant du côté droit de la rétine et ce, pour les deux yeux (champ visuel gauche). • Colliculi supérieurs : Structure souscorticale. Reçoit environ 10% des fibres ganglionnaires. – Impliqué dans le contrôle des mouvements des yeux.
Traitement de l’information dans le CGL Du cortex Vers cortex Thalamus-LGN 10 axones de la rétine 4 axones vers le cortex Tronc cérébral De la rétine + En plus de recevoir les signaux optiques, le CGL reçoit des entrées en provenance du cortex visuel, du tronc cérébral, d’autres noyaux thalamiques et d’autres neurones du CGL. = Jouerait un rôle dans la régulation de l’information en provenance de la rétine.
Les champs récepteurs du CGL Le champs récepteur d’un neurone du CGL est une région sur la rétine qui, lorsque stimulée, a un effet maximal sur le déclenchement de ce neurone. = luminance haute Le fond de la rétine = luminance basse = peu importe
Champs récepteur d’une cellule du LGN (“center-surround”) • Les champs récepteurs des neurones du CGL ont la même configuration concentrique les cellules ganglionnaires. Il existe aussi des champs récepteurs “center OFF-surround ON” dans le LGN. (Ozhawa, De. Angelis & Freeman 1995)
Organisation dans le LGN (couches et oeil d’origine) Chaque LGN a 6 couches (la couche 1 étant la plus profonde et la 6 la plus superficielle). Coupe coronale Oeil ipsilatéral : couches 2, 3 et 5 Oeil controlatéral : couches 1, 4 et 6
Organisation dans le LGN (rétinotopie) Chaque couche d’un LGN est rétinotopique. Les neurones situés près les uns des autres dans une structure cérébrale ont des champs récepteurs correspondant à des localisations rétiniennes voisines. Oeil
Organisation dans le LGN (magno vs parvo) • Les couches 1 et 2 sont magnocellulaires. C’est-à-dire qu’elles reçoivent des axones de cellules ganglionnaires de type M). • Les autres couches (3, 4, 5, 6) sont parvocellulaires. C’est-à-dire qu’elles reçoivent des axones de cellules ganglionnaires de type P).
Organisation dans le LGN (magno vs parvo II) • Les cellules des couches magnocellulaires et parvocellulaires ont des rôles différents dans la perception visuelle. • Lésions des couches magno : – Atteinte de la perception du mouvement. • Lésions des couches parvo : – Atteinte de la perception des couleurs, de la texture, de la profondeur et des contours.
Plan pour le reste du cours Corps genouillé latéral (LGN) droit lumière Corps genouillé latéral (LGN) gauche Cortex visuel primaire (V 1)
Régions de traitement du signal visuel dans le cortex Cortex visuel primaire (V 1) ou cortex strié Cortex extrastrié
Régions de traitement du signal visuel dans le cortex Cortex visuel primaire (V 1) ou cortex strié Cortex extrastrié
Cortex strié Contient environ 200 millions de neurones, plus de 100 fois plus que le CGL (environ 1. 5 millions).
D. Hubel T. Wiesel Nobel en physiologie en 1981
Cortex visuel primaire • Au contraire des neurones du CGL, ceux du cortex strié ne répondent pas particulièrement bien à des points lumineux.
Ozhawa, De. Angelis & Freeman 1995 Classique Corrélation inversée http: //www. youtube. com/watch? v=KE 952 yue. VLA http: //www. youtube. com/watch? v=QZUzeny 0 s. Rg&feature=related
Origine des cellules simples?
Le rôle des cellules simples Phase Orientation Rappelez-vous : onde sinusoidale = Amplitude * sin(fréquence * angle + phase) Ici on ajoute l’orientation Contraste Fréquence
Cellules simples vs complexes • • • Les cellules simples ne répondent qu’à des stimuli ayant la bonne orientation et le bon positionnement dans leur champ récepteur. Les cellules complexes sont également sélectives à l’orientation mais répondent quelle que soit la position du stimulus dans leur champ récepteur. En moyenne, les cellules complexes tendent à avoir des champs récepteurs plus étendus que les cellules simples.
Cellules hypercomplexes • La réponse des cellules hypercomplexes (ou à inhibition terminale) varie en fonction de la longueur du stimulus.
Propriétés des neurones du nerf optique, du CGL et du cortex
Organisation dans V 1
Rétinotopie dans V 1 révélée par autoradiographie • On montre cette image à un singe + injection de sucre radioactif.
Rétinotopie dans V 1 : Résultats Stimulus Cortex
Propriétés du cortex strié: Magnification corticale • La surface corticale dédiée à la représentation d’un stimulus sur la rétine dépend de sa localisation sur la rétine. • Une plus grande surface du cortex visuel primaire est ainsi dédiée au traitement de la fovéa par rapport à la périphérie. • Ainsi la fovéa correspond à environ 0. 01% de la rétine alors que 8 à 10% du cortex visuel primaire est dédié au traitement de l’information fovéale.
Magnification corticale • En partie responsable du fait que l’acuité visuelle décline avec l’excentricité. • Mais pourquoi plus de ressources corticales au centre que dans la périphérie? – Une haute résolution spatiale nécessite: • Plus de photorécepteurs dans la fovéa par unité de surface. • Une relation un à un entre les photorécepteurs et les cellules ganglionnaires. • Plus grand nombre de cellules corticales dédiées à chaque cellule de la fovéa. Notre cerveau serait trop gros si les mêmes ressources étaient dédiées à la périphérie
Rétinotopie dans V 1 : Résultats
Dominance oculaire • V 1 est la première aire visuelle à avoir des neurones qui reçoivent de l’information des deux yeux. – 80% des neurones du cortex strié ont des champs récepteurs binoculaires (i. e. pour être stimulé par un œil ou l’autre). – Les 20% restants sont monoculaires. • Les neurones ayant la même dominance oculaire sont organisés en colonne.
Colonnes d’orientation • Lorsqu’une électrode suit une trajectoire perpendiculaire, les neurones ont la même préférence en terme d’orientation (et la même dominance oculaire).
Colonnes d’orientation (suite) • Lorsqu’une électrode suit une trajectoire oblique, les champs récepteurs des neurones rencontrés présentent une variation continue de l’orientation préférée. • Une progression de 1 mm permet de traverser toute l’étendu possible des orientations pour les deux types de dominance oculaire.
Sélectivité à l’orientation • Le neurone s’active davantage pour une barre verticale. • Le neurone ne réagit plus du tout à +/- 30 deg. • Le cortex visuel contient des neurones dont le stimulus favori est à différentes orientations. Ainsi, toutes les orientations possibles peuvent être détectées.
Les hypercolonnes Une hypercolonne mesure environ 1 mm 3; et “voit” entre 3 min d’angle visuel et 0. 7 deg d’angle visuel. perpendiculaire 1 hypercolonne = 2 colonnes de dominance oculaire = 2 ensembles des colonnes d’orientation oblique
Rétinotopie • L’ensemble des hypercolonnes est rétinotopique (c’est-àdire que 2 hypercolonnes adjacentes ont des champs récepteurs adjacents sur les rétines). Mais il y a grossissement à la fovéa.
Le rôle des cellules simples Phase Orientation Contraste Fréquence
Relation avec les cellules simples • En fait, les cellules simples sont sensibles à trois attributs des stimulations rétiniennes – La fréquence spatiale. – l’orientation. – et la position sur la rétine. Démonstration
Adaptation • Exposition continue, pour une certaine période de temps, à un stimulus comportant une propriété spécifique. • L’effet d’adaptation se manifeste par un effet consécutif et est habituellement expliqué par une fatigue cellulaire sélective. – Effet consécutif : Modification du fonctionnement perceptif suite à l’exposition prolongée à une stimulation.
Adaptation • Si un stimulus (ou une propriété donnée d’un stimulus) provoque un effet consécutif, c’est que ce stimulus (ou cette propriété) est 1) traité(e) par le système perceptif 2) les neurones associés ont un effet direct sur la perception.
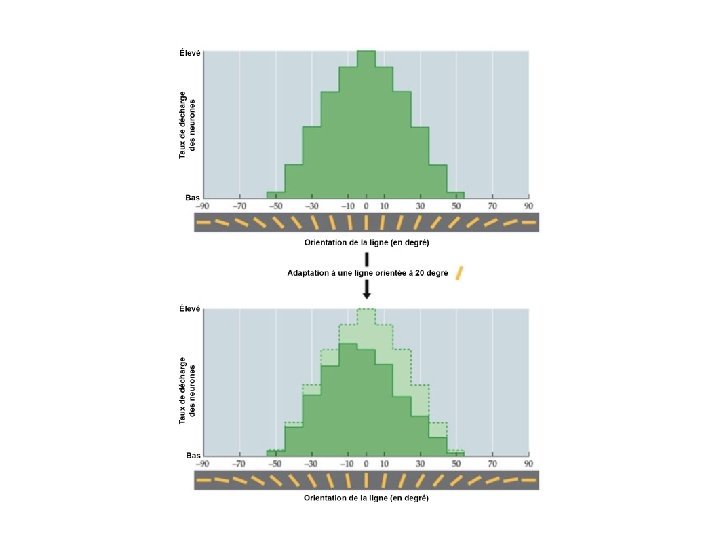
Adaptation à l’orientation *
Adaptation à l’orientation *
Adaptation à l’orientation *
Adaptation à la fréquence spatiale *
Adaptation aux fréquences spatiales *
Adaptation aux fréquences spatiales *
Sensibilité au contraste • Niveau de contraste minimal pour détecter l’alternance entre les barres pâles et foncées constituant une grille sinusoïdale. Contraste élevé Contraste moyen Contraste faible
Approximation de votre courbe de sensibilité au contraste
Courbe de sensibilité au contraste Le système visuelle humain a moins de neurones pour les basses fréquences spatiales 60 cycles par deg Résolution visue limite chez l’hum
Que font les hypercolonnes?
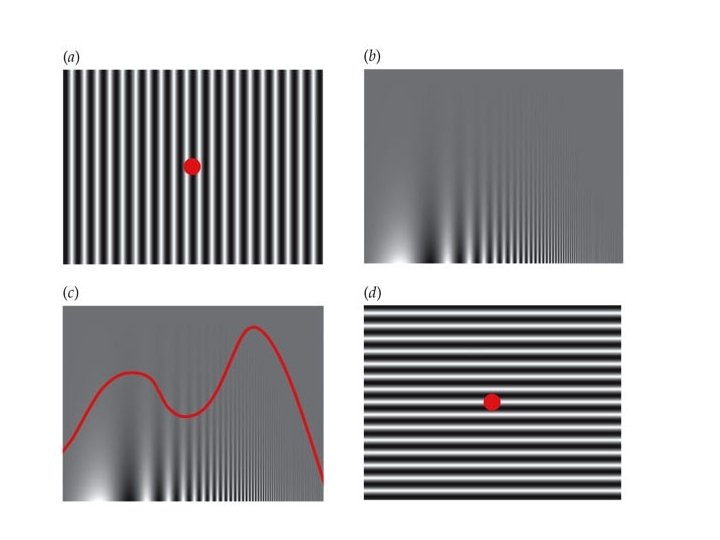
Une analyse de Fourier (locale)! = + réseau Analyse de Fourier 2 D : une décomposition unique d’une image en une somme de grilles sinusoïdales.
Une analyse de Fourier (locale)! = + Fréquence = 1 + + Fréquence = 5 Fréquence = 3 + … + Fréquence = 7 Fréquence = 9
Une analyse de Fourier (locale)!
(Adapté de Schyns et Oliva, 1999)
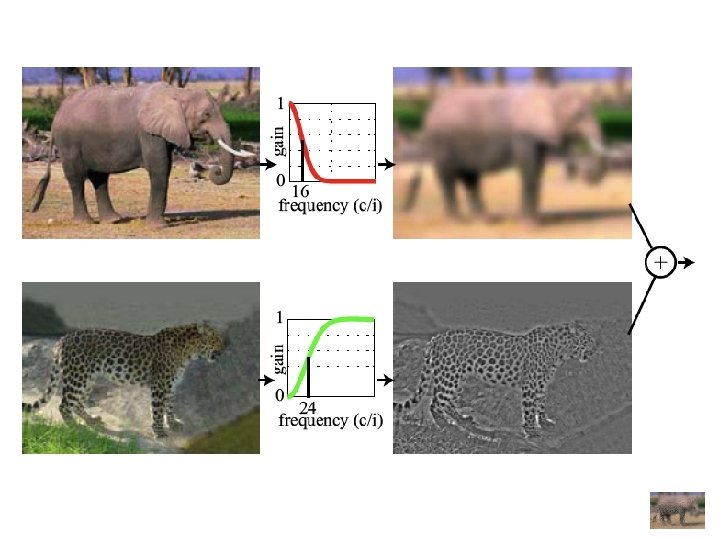
Filtrage et fabrication d’un hybride Filtre “passe-haute” Combinaison Filtre “passe-basse”
Hybride
Retour au pseudo-paradoxe de la perception • Si le monde est tel que nous le percevons, le cerveau est tel que nous le percevons; • Or 50% de la lumière visible ambiante traverse le médium oculaire * 20% traverse les cellules de la rétine * moins de 1% de l’information dans les récepteurs rétiniens sort des cellules ganglionnaires * 40% de l’information qui arrive dans le LGN le quitte pour V 1 = moins de 0, 04% de l’information dans la lumière visible parvient au cortex extrastrié; • Donc le monde n’est pas tel que nous le percevons.