PROTEN SENTEZ ve POSTTRANSLASYONEL MODFKASYONLAR Santral Doma n








")
 1. Aminoasitlerin aktivasyonu: 20 aminoacyl-t. RNA sentezlenmesi, 32")






 eklenmesi (proteinin zara bağlanması")

 n Genellikle C-terminali yakınındaki sisteinlerin kükürt atomuna izoprenoid lipidler (farnesil")
 Açilasyon, C veya N-terminalinde olabilir. En sık miristik asit ile")









 bağlanması n Enzimlerin inaktif apo formları, aktif holo formlarına çevrilir. n")
- Slides: 31
PROTEİN SENTEZİ ve POSTTRANSLASYONEL MODİFİKASYONLAR
Santral Doğma n DNA- DNA Replikasyon n DNA- RNA Transkripsiyon n RNA- Protein Translasyon
PROTEİN SENTEZİ Protein sentezi hücrede ribozamlarda gerçekleşir. Ribozomlar protein ve r. RNA’dan oluşmaktadır. Bakteri ribozomu 70 S (50 S ve 30 S), ökaryot hücre ribozomu 80 S subunitine ve daha fazla proteine sahiptir. n Protein sentezinde 3 Tip RNA görevli r. RNA: En fazla bulunan RNA. En uzun ömürlüdür. m. RNA: En az miktardadır. En kısa ömürlüdür t. RNA: En küçükleridir. Akseptör kolu, Antikodon kolu, D kolu (Dihidroüridin taşır ve uygun aminoaçil-t. RNA sentetazın tanınması), TψC kolu (thymidine, pseudouridine cytosine taşır ve ribozom üzerine uygun yere bağlanma), Ekstra kol n
Bazı Önemli Tanımlamalar n n Genlerdeki ve dolayısıyla m. RNA’daki nükleotid dizisi ile proteinlerdeki amino asit dizisi arasındaki ilişki genetik şifre’dir. Nükleotidlerin m. RNA üzerindeki üçlü dizilimlerine kodon adı verilir. Her bir kodon bir amino asidi şifreler. Başlama kodonu: N-Formilmethionin t. RNA’nın tanıdığı AUG kodonu (Metionini kodlayan kodon) Stop kodonu: (UAA, UAG ve UGA) Protein sentezini durduran kodonlardır. Herhangi bir a. a. ’i kodlamazlar
n Protein sentezinde başlangıç a. a. ’i metionindir, kodonu ise AUG’dir. n Metionini 2 t. RNA’sı vardır biri peptid başlatıcı t. RNA (N-formil metionil t. RNA) diğeri peptid yapı içerisine yerleştirilecek metionin için olan peptid metionin t. RNA.
n Protein sentezi metionin a. a’inin m. RNA’daki kodonu olan AUG kodonunun ve sıradaki diğer a. a’lerin kodonlarının t. RNA’nın akseptör ucundaki CCA 3’ terminalindeki Adenozin bazına bağlanmasıyla başlar. Her hücrede her bir aminoasite özgü bulunan aminoaçil t. RNA sentetaz enzimi t. RNA’nın D koluna bağlanır ve t. RNA ile a. a arasında bağ oluşturur.
n Aminoaçil t. RNA sentetaz açilasyon için doğru a. a. seçimini ve amino-açillenmiş t. RNA doğru a. a. alıp almadığını defalarca kontrol eder.
n m. RNA kodonu 5’ ucundan 3’ ucuna doğrudur n Protein sentezi a. a’lerin amino grubundan karboksil grubu yönünedir. n Bu nedenle m. RNA’nın 5’ ucu sentezlenen proteinin amino ucuna, 3’ ucu ise karboksil ucuna gelecek şekilde pozisyon alır.
Protein Sentez Basamakları 5 tanedir n Aminoasitlerin Aktivasyonu n Başlangıç basamağı n Uzama (Elongasyon) basamağı n Sonlanma basamağı n Posttranslasyonel Modifikasyon basamağı
Protein sentezinde gerekli komponentler (E. Coli) 1. Aminoasitlerin aktivasyonu: 20 aminoacyl-t. RNA sentezlenmesi, 32 veya daha çok t. RNA, ATP ve Mg 2. Başlama basamağı: m. RNA, N-Formylmethionylt. RNAfmet, Başlama kodonu m. RNA (AUG), 30 S ribozomal subunit, 50 S ribozomal subunit, Başlama faktörleri (IF-1, IF-2, IF-3), GTP ve Mg 3. Uzama basamağı (Translokasyon): Fonksiyonel 70 S ribosome (başlama kompleksi), Aminoaçil-t. RNA’lar , Elangasyon faktörleri (EF-Tu, EF-Ts, EF-G), GTP ve Mg 4. Sonlanma ve salınım: Sonlanma kodonu (m. RNA’da), Release faktörler (RF-1, RF-2, RF-3) 5. Katlanma ve posttranslasyonel süreç: Spesifik enzimler, kofaktörler, eklenecek çeşitli gruplar (phosphate, methyl, carboxyl, carbohydrate)
Protein Sentez İnhibitörleri n Kloramfenikol: Prokaryotlarda peptidil transferaz inhibisyonu yapar n Streptomisin, Neomisin: Prokaryotlarda m. RNA’nın yanlış okunmasını sağlayarak peptid oluşumunu engeller. n Tetrasiklin: Prokaryotlarda aminoaçilt. RNA’nın ribozama bağlanmasını engelller. n Eritromisin: Prokaryotlarda translokasyonu inhibe eder.
n n n Fusidik asit: EF-disasyonunu engelleyerek translokasyona engel olur. Promisin: Aminoaçil-t. RNA analoğudur. Hem ökaryot hem de prokaryotlarda peptid zincire katılır fakat translokasyon gerçekleşemez. Difteri toksini: Ökaryotlarda EF 2 elongasyon faktörünü inhibe eder. Risin: Ökaryotlarda ribozom inaktive edici protein olarak da bilinir. Biyolojik silah olarak da kullanılmıştır. Siklohekzamid: Ökaryotlarda peptidil transferaz enzimini inhibe eder.
POSTTRANSLASYONEL MODİFİKASYONLAR
PTM; Proteinlerin yan zincirlerinde translasyonu ve ribozomdan çıkışı takiben meydana gelen enzimatik kovalent modifikasyonlardır. AMAÇ: Proteinlerdeki fonksiyonel grupları 20 amino asitin ötesinde çeşitlendirerek proteinin ya fonksiyonunu ya da lokalizasyonunu değiştirmektir.
PTM sonucunda; n Protein katlanmasında değişiklik yapılabilir. n Proteinler yıkım için işaretlenebilir. n Kromatinde transkripsiyonel bölgeler işaretlenebilir. n Proteinlerin intra ve ekstrasellüler adresleri değiştirilebilir. n Enzimler inaktif iken aktif formlarına çevrilebilir.
PTM’ların Hücresel Yerleşimleri Sitoplazmadakiler: n Başlatıcı Met’in çıkarılması n N-terminalinde asetilasyon n O-glikozilasyon n Palmitoil gruplarının eklenmesi
RER’da: n Sinyal peptid ayrılması n Palmitoil ve glikozilfosfatidilinozitol (GPI) eklenmesi (proteinin zara bağlanması için) n Hidroksilasyon, glikozilasyon n Disülfid bağı oluşumu (tersiyerkuarterner yapı) Golgi’de: n N-glikozil gruplarının modifikasyonu n O-glikozilasyon n Tirozinlerin sülfatlanması n Proteolitik işleme (prohormon)
Glikozilasyon Hücre zarında bulunan glikoproteinler, müköz zarları kaplayan kayganlaştırıcı proteoglikanlar ve bazı hormonlar karbonhidrat yan zinciri taşır. n N-glikozilasyon genellikle Asn üzerinden kotranslasyonel olarak yapılır. Dolikol fosfat ER’da karbonhidratları taşıyan lipidsi yapıdır. n n O-glikozilasyon genellikle Ser ve Thr’in -OH grupları üzerinden ve başlıca Golgi’de posttranslasyonel olarak gerçekleşir. n Eklenecek glukoz UDP ile, mannoz GDP ile aktive edilir.
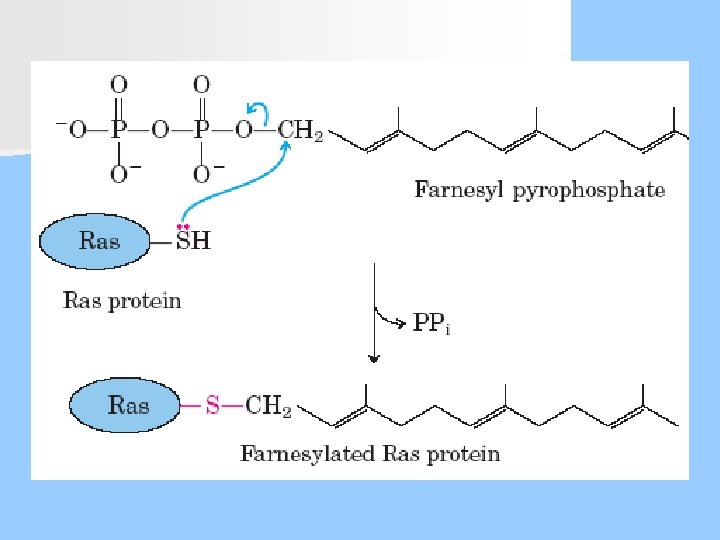
Prenilasyon (İzoprenil grupları eklenmesi) n Genellikle C-terminali yakınındaki sisteinlerin kükürt atomuna izoprenoid lipidler (farnesil ve geranil) eklenir. n Prenillenmiş proteinlere örnek olarak ras onkogeni, görmede görev yapan transdusin ve G-proteinleri verilebilir. n Bu izoprenil grupları molekülü zara gönderip, orada demirlenmesini sağlar.
Yağ asidi eklenmesi (Açilasyon) Açilasyon, C veya N-terminalinde olabilir. En sık miristik asit ile palmitoik asit eklenir. n Genellikle intrensek zar proteinleri bu yolla modifiye edilir. Bu ekleme proteini zara yönlendirir. n Ras onkogeninin C-terminaline palmitik asit eklenir. c. AMP bağımlı protein kinazlar ile tirozin kinazların katalitik alt ünitesine, G proteinlerinin alt ünitesine de miristik (C 14) asit eklenir. n
Asetilasyon n Histonlarda bulunan Lys bakiyelerine asetiltransferaz enzimleriyle asetil grubu eklenir. Bu işlem histonların transkripsiyonel aktivasyonu, kromatin organizasyonu ve DNA replikasyonu için gereklidir.
Metilasyon n Proteinlerin Lys bakiyelerine SAM aracılığıyla metil eklenmesi bakteriyel kemotaksis için gerekli olabilir. n Örnek: Kalmodulin ve sitokrom C’nin Lys bakiyelerine yine SAM aracılığıyla metilasyon yapılır.
Hidroksilasyon n Kollajendeki Pro ve Lys bakiyelerinin hidroksilasyonu, C vitamini koenzim olarak kullanılanarak prolil ve lizil hidroksilaz tarafından yapılır. Bu, takiben gerçekleşecek olan glikozilasyon ve tropokollajen zincirlerinin birbirlerine sarılması ve kollajen lifçiği oluşturulması için gerekli bir modifikasyondur.
Fosforilasyon En sık görülen modifikasyondur. n Bir proteinin biyolojik aktivitesini değiştirmek için yapılır, bu yüzden geçicidir. n Genellikle Ser, Thr, Tyr ve Cys bakiyelerinin – OH / -SH grupları ATP kullanılarak kinaz enzimleriyle fosforlanır: ATP + protein <----> fosfoprotein + ADP n Fosforilazlarla da defosforile edilir. n En tipik örnek metabolik enzimlerdir: ör. Karaciğerdeki glikojen sentaz ve glikojen fosforilazın fosforlanması. n
Sülfat eklenmesi n Bazı proteinlerin tirozin bakiyelerine biyolojik aktiviteyi sağlamak için sülfat eklenmesi gerekir. Ör: gastrin, fibrinojen. n Sülfat vericisi 3'-fosfoadenozil-5'fosfosülfat (PAPS) tır.
Karboksilasyon n Pıhtılaşma faktörleri, ör: protrombinin Nterminalindeki Glu bakiyelerinin karboksilasyonu, K vitamini koenzim olarak kullanılarak yapılır. n Bu –COO grupları Ca++ ile bağlanmayı sağlar. n Karboksilasyon ile -karboksiglutamat oluşur:
Proteolitik İşleme n n n Birçok protein translasyondan sonra proteolize uğrar. Bunun en basit örneği başlatıcı Met’in çıkarılmasıdır. Pek çok protein belli fizyolojik koşullarda aktiflenmek üzere inaktif olarak üretilir. Sekresyon sonrasında proteolize uğrayarak aktiflenen inaktif öncü proteinlere proprotein denir. Ör: pankreatik sindirim enzimleri (zimojenler). Preproprotein denilen bazı proteinler de hem sekresyon öncesi hem de sonrasında iki kez proteolize uğrar, ilk ayrılan pre kısmına sinyal dizi de denir, ör: hormonlar (insülin).
Disülfid bağı kurulması ve protein katlanması n n n Disülfid bağı Cys’lerdeki –SH gruplarının oksidatif bağlanmasıyla kurulur. RER’da olur. Tersiyer ve kuarterner yapıyı stabilize eder, normal aktivite için gereklidir. Sekretuar proteinlerin katlanması sıklıkla RER’da olur, bu işleme yardımcı olan proteinler vardır, ör: şaperonlar. Kuarterner yapı kurulması da genellikle Golgi’ye gönderilmeden az önce olur, yine şaperonların yardımıyla. Yalnız düzgün katlanmış proteinler RER’dan Golgi’ye taşınabilir, aksi halde içeride kalarak yıkılır.
Ubikitin bağlanması n Kendisi de 76 bakiyelik küçük bir protein olan ubikitin, hedefteki proteinin Lys yan zincirlerine bağlanır. n Ubikitinlenmiş protein, proteazomlar tarafından tanınarak proteolitik yıkıma gider. n RER’da şaperonlara bağlı olarak bekletilen hatalı katlanmış veya katlanamamış proteinler ile ömrü dolmuş proteinler, ubikitin-proteazom sisteminde yıkılırlar.
Koenzim (prostetik grup) bağlanması n Enzimlerin inaktif apo formları, aktif holo formlarına çevrilir. n Enzim proteini ribozomdan çıktıktan sonra bağlanma olur. n Ör: yağ asidi sentaz enzimine fosfopantetein, karboksilazlara biotin, piruvat dehidrogenaza lipoik asit eklenmesi.