Protein synthesis I Genetic code The genetic code


 can")
 codons: • Three of the codons, UAG, UGA, and UAA,")


















- Slides: 24
Protein synthesis
I. Genetic code ü The genetic code is the way in which the nucleotide sequence in m. RNA ( or DNA ) specifies the amino acid sequence in protein. ü Their nucleotide sequences are always written from the 5'-end to the 3'-end. ü The four nucleotide bases are used to produce three-base codons. There are, therefore, 64 different combinations of bases, taken three at a time.
How to translate a codon • • The genetic code table (or “dictionary”) can be used to translate any codon and, thus, to determine which amino acids are coded for by an m. RNA sequence. For example, the codon 5' -AUG-3' codes for methionine Sixty-one of the 64 codons code for the 20 common amino acids.
Termination (“stop” or “nonsense”) codons: • Three of the codons, UAG, UGA, and UAA, do not code for amino acids, but rather are termination codons. • When one of these codons appears in an m. RNA sequence, synthesis of the polypeptide coded for by that m. RNA stops.
Characteristics of the genetic code 1. 2. 3. 4. Specificity. Universal. Degeneracy. Nonoverlapping and commaless.
Consequences of altering the nucleotide sequence: q Changing a single nucleotide base on the m. RNA chain (a “point mutation”) can lead to any one of three results: 1. Silent mutation: 2. Missense mutation: 3. Nonsense mutation:
II. Components required for translation
A. Amino acids: All the amino acids that eventually appear in the finished protein must be present at the time of protein synthesis B. Transfer RNA: At least one specific type of t. RNA is required for each amino acid. 1. Amino acid attachment site: Each t. RNA molecule has an attachment site for a specific (cognate) amino acid at its 3'-end. 2. Anticodon: Each t. RNA molecule also contains a three-base nucleotide sequence—the anticodon— that pairs with a specific codon on the m. RNA.
How many t. RNA molecules are there? Wobble rules First Base of Anticodon Pairs with: Third Base of codon Normal By wobble G C Or U U A Or U I !!!! - C or U or A C G only No wobble A U only No wobble Unusual base
C. Aminoacyl-t. RNA synthetases : This family of enzymes is required for attachment of amino acids to their corresponding t. RNAs. . The extreme specificity of the synthetase in recognizing both the amino acid and its cognate t. RNA contributes to the high fidelity of translation of the genetic message. In addition, the synthetases have a “proofreading” or “editing” activity that can remove amino acids from the enzyme or the t. RNA molecule. D. Messenger RNA: The specific m. RNA required as a template for the synthesis of the desired polypeptide chain must be present.
E. Functionally competent ribosomes : Ribosomes are large complexes of protein and ribosomal RNA They consist of two subunits one large and one. The small ribosomal subunit binds m. RNA and is responsible for the accuracy of translation by ensuring correct base-pairing between the codon in the m. RNA and the anticodon of the t. RNA. The large ribosomal subunit catalyzes formation of the peptide bonds that link amino acid residues in a protein.
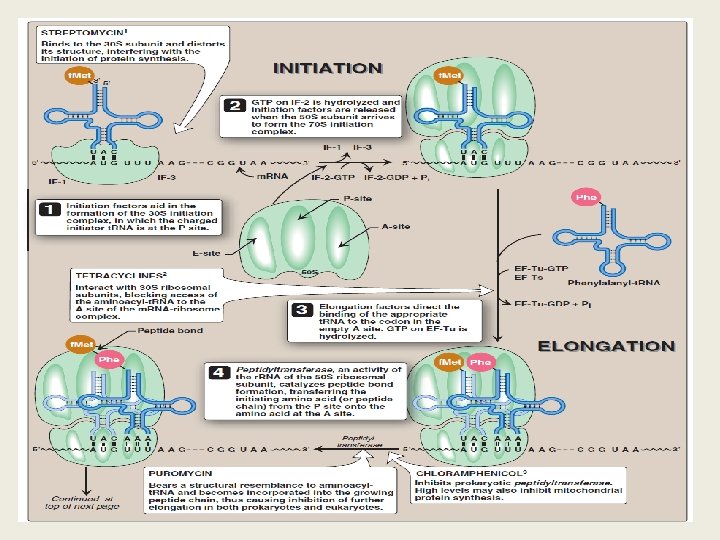
III. STEPS IN PROTEIN SYNTHESIS • • • The process of protein synthesis translates the three-letter alphabet of nucleotide sequences on m. RNA into the 20 -letter alphabet of amino acids that constitute proteins. The m. RNA is translated from its 5'-end to its 3'-end, producing a protein synthesized from its amino-terminal end to its carboxyl terminal end. Prokaryotic m. RNAs often have several coding regions, that is, they are polycistronic. Each coding region has its own initiation and termination codon and produces a separate species of polypeptide. In contrast, each eukaryotic m. RNA has only one coding region, that is, it is monocistronic. The process of translation is divided into three separate steps: initiation, elongation, and termination.
• • One important difference is that translation and transcription are coupled in prokaryotes, with translation starting before transcription is completed. Coupling is a consequence of the lack of a nuclear membrane in prokaryotes.
Shine-Dalgarno sequence Complementary binding between prokaryotic m. RNA Shine-Dalgarno sequence and 16 S r. RNA. o In E. coli, a purine-rich sequence of nucleotide bases, is located six to ten bases upstream of the initiating AUG codon on the m. RNA molecule—that is, near its 5'-end. o Eukaryotic messages do not have SD sequences. o In eukaryotes, the 40 S ribosomal subunit binds close to the cap structure at the 5 -end of the m. RNA and moves down the m. RNA until it encounters the initiator AUG.
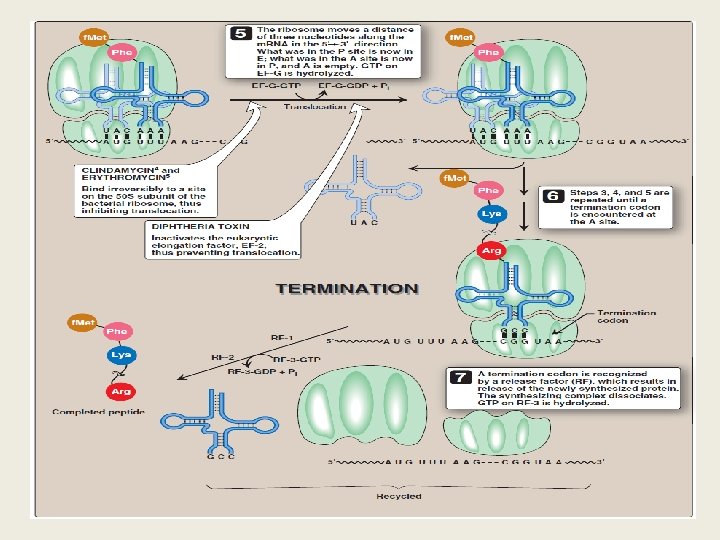
B- Elongation Ø Elongation of the polypeptide chain involves the addition of amino acids to the carboxyl end of the growing chain. Ø Delivery of the aminoacyl-t. RNA whose codon appears next on the m. RNA template in the ribosomal A site is facilitated in E. coli by elongation factors EF-Tu-GTP and EF-Ts, and requires GTP hydrolysis. Ø After the peptide bond has been formed, what was attached to the t. RNA at the P site is now linked to the amino acid on the t. RNA at the A site. Ø The ribosome then advances three nucleotides toward the 3'-end of the m. RNA. This process is known as translocation. Ø Translocation causes movement of the uncharged t. RNA from the P to the E site, and movement of the peptidyl -t. RNA from the A to the P site. Ø The process is repeated until a termination codon is encountered.
Formation of a peptide bond
C- Termination Ø Termination occurs when one of the three termination codons moves into the A site. These codons are recognized in E. coli by release factors: RF-1, which recognizes the termination codons UAA and UAG, and RF-2, which recognizes UGA and UAA. Ø The binding of these release factors results in hydrolysis of the bond linking the peptide to the t. RNA at the P site, causing the nascent protein to be released from the ribosome. Ø A third release factor, RF-3 -GTP then causes the release of RF-1 or RF-2 as GTP is hydrolyzed.
An overview of the main events in translation. Translation is initiated by the pairing of an m. RNA and t. RNA on the ribosome. In elongation, the ribosome moves along the m. RNA, matching t. RNAs to each codon and catalyzing peptide bond formation. Translation terminates at a stop codon, and the ribosomal subunits are released for another round of synthesis
III. CO- and Posttranslational modification of polypeptide chains Ø Many polypeptide chains are covalently modified, either while they are still attached to the ribosome (cotranslational) or after their synthesis has been completed (posttranslational). Ø These modifications may include removal of part of the translated sequence, or the covalent addition of one or more chemical groups required for protein activity.
Ø Some types of posttranslational modifications are listed below. A. Trimming Many proteins destined for secretion from the cell are initially made as large, precursor molecules that are not functionally active. Portions of the protein chain must be removed by specialized endoproteases, resulting in the release of an active molecule. The cellular site of the cleavage reaction depends on the protein to be modified. • • • B. Covalent attachments • Proteins may be activated or inactivated by the covalent attachment of a variety of chemical groups. • Phosphorylation • Glycosylation • Hydroxylation
C. Protein folding • Proteins must fold to assume their functional state. Folding can be spontaneous (as a result of the primary structure), or facilitated by proteins known as “chaperones”. D. Protein degradation • • Proteins that are defective, for example, misfolded, or destined for rapid turnover are often marked for destruction by ubiquitination— the attachment of chains of a small, highly conserved protein, called ubiquitin. Proteins marked in this way are rapidly degraded by a cellular component known as the proteasome, which is a macromolecular, ATP-dependent, proteolytic system located in the cytosol.