POTENCIALES BIOELECTRICOS FISIOLOGA I 2 DO CICLO TEMA

POTENCIALES BIOELECTRICOS FISIOLOGÍA I, 2 DO. CICLO, TEMA 3 LICENCIATURA EN HOMEOPATÍA

INTRODUCCIÓN Un campo eléctrico es un campo de fuerza creado por la atracción y repulsión de cargas eléctricas. Su aplicación en el tejido biológico fue iniciada por el fisiólogo alemán Emil Du Bois-Reymond (1818 -1896) quien documentó en detalle las actividades eléctricas asociadas a la excitación nerviosa, contracción muscular y heridas, llevándolo a ser uno de los fundadores de la electrofisiología moderna. Los campos eléctricos (CE) se encuentran de manera natural en los organismos vivos y son encargados de controlar procesos de crecimiento y comportamientos celulares.

POTENCIAL DE ACCIÓN Los potenciales de acción son cambios del potencial de membrana que se propagan a lo largo de la superficie de células excitables. Los potenciales bioeléctricos o biopotenciales son voltajes iónicos producidos como resultado de la actividad electroquímica de ciertos tipos especiales de células denominadas excitables, que pueden ser nerviosas, musculares y glandulares. “El potencial de membrana” es la energía eléctrica almacenada como voltaje, por las pequeñas baterías que son células. Esta energía se utiliza en muchas funciones celulares.

La vista, el oído, las señales de dolor son transmitidas a través de impulsos eléctricos que se transmiten a través de los nervios. Al colocar electrodos sobre la piel se puede registrar los potenciales eléctricos generados por esa corriente. Un ejemplo de estos trazos son el electroencefalograma, la electromiografía y el electrocardiograma. Cada tipo de célula tiene una actividad eléctrica característica, por lo tanto las señales bioeléctricas tienen una amplitud y forma distinta.

Los líquidos orgánicos son soluciones conductoras que contienen átomos cargados llamados iones. Se denomina electrón si la carga mínima o carga elemental es negativa y se denomina protón, si es positiva. Los principales son el sodio, el potasio y el cloro. Los espacios intra y extracelular están separadas por un material aislante o dieléctrico, la membrana celular, que es una bicapa fosfolipídica. Por lo tanto, podemos ver células como pequeñas baterías, que guardan una diferencia de potencial o voltaje (diferencia de carga) entre su interior y el exterior. En proporción hay más cargas negativas en el interior celular.
, disueltos")
La membrana lipídica de la célula separa carga eléctrica. Los iones cargados (electrólitos), disueltos en los líquidos intracelulares y extracelulares, están separados de manera asimétrica por la membrana celular. Hay más cargas negativas (aniones) que positivas (cationes) en el interior celular, lo que genera un voltaje transmembranal. Energía eléctrica almacenada lista para ser utilizada cuando la célula la necesite. Tal como en una batería o pila.
 es")
LEY DEL TODO O NADA Una importante propiedad del Potencial de Acción (PA) es que sigue la ley del todo o nada, según la cual, cuando un estímulo (o una serie de estímulos) llega a la neurona con suficiente intensidad como para provocar la aparición de un PA en la membrana de dicha célula, el PA se produce en todo caso, y alcanzando la amplitud (voltaje) máxima posible en cada caso (ese máximo no es de idéntica magnitud en cada caso, pero será la magnitud máxima posible en cada suceso). No por aumentar la intensidad del estímulo aumenta también el potencial de acción, no existen respuestas intermedias, todo o nada. Si se imagina que el potencial de acción fuera una flecha disparada con una ballesta de una neurona a otra, siguiendo la ley del todo o nada (o se dispara, o no se dispara, sin término medio), se puede imaginar que el todo o nada se prepara en el instante previo cargando la ballesta.

Si un estímulo ha sido tan intenso como para que la despolarización que provoca cruce el umbral de aparición del PA, éste aparece y el potencial bioeléctrico se propaga sin que la magnitud de dicha amplitud se resienta, hasta llegar a la siguiente estación de relevo del flujo eléctrico, la sinapsis, desde donde se suelta una cantidad de neurotransmisor proporcional a la amplitud del PA, de modo que a la membrana postsináptica llega suficiente neurotransmisor como para desencadenar otro PA (sobre todo, si son varias las neuronas convergentes en la región postsináptica). Una vez que se genera este potencial de acción tiene la misma duración y amplitud que si se inicia por un estímulo umbral o supraumbral.

UMBRAL DE EXCITACIÓN Este concepto se refiere a la intensidad mínima que debe tener un estímulo para ser capaz de generar un potencial de acción en la neurona. De acuerdo a esto existen tres clases de estímulos según su intensidad: 1. Estímulo umbral, es aquel que tiene la intensidad mínima necesaria para generar un potencial de acción. 2. Estímulo subumbral (Infraumbral), que tiene una intensidad menor al mínimo necesario y por tanto no es capaz de generar potencial de acción. 3. Estímulo supraumbral, tiene una intensidad mayor al mínimo necesario y también es capaz de generar potencial de acción

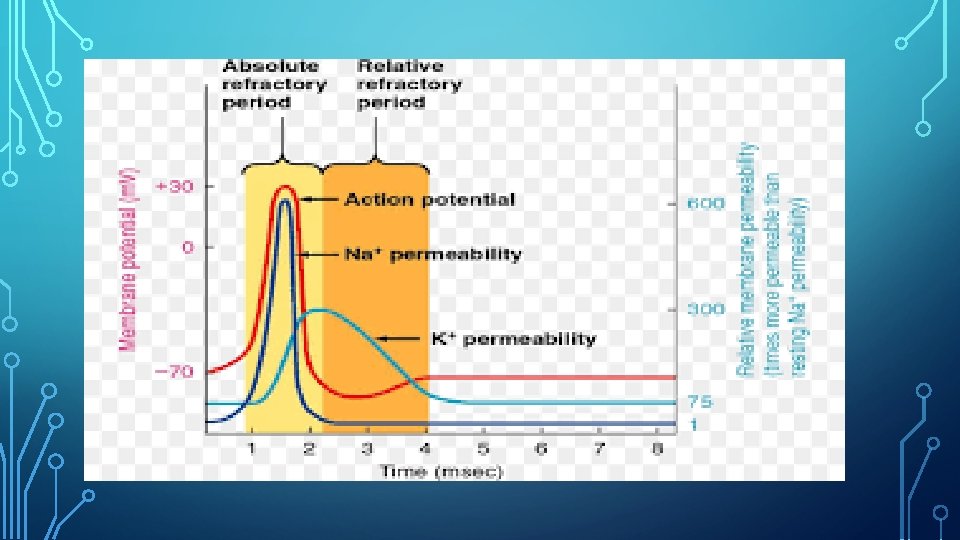
PERIODO REFRACTARIO Es el periodo comprendido entre el inicio de dla despolarización y el final de la repolarización. Se divide en absoluto y relativo. Se refiere a un lapso en el cual la neurona no puede volver a ser excitada y ocurre durante la repolarización. En un principio el período refractario es absoluto, porque la neurona no responde a ninguna clase de estímulo incluso los supraumbrales; para luego volverse relativo en donde sólo responde a estímulos supraumbral.

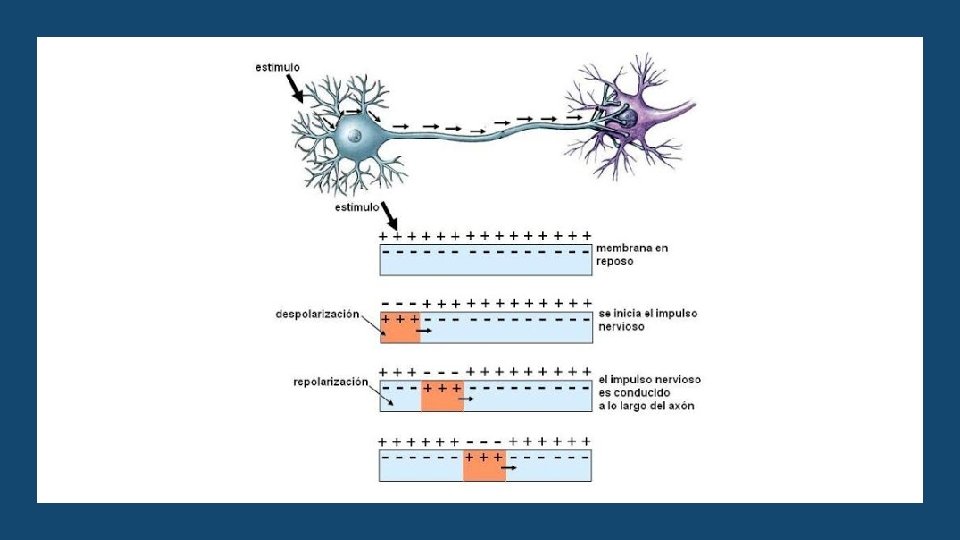
CONDUCCIÓN DE IMPULSO En las neuronas que carecen de vaina de mielina se habla de una condición continua porque el potencial de acción va despolarizando toda la membrana neuronal. Sin embargo, las neuronas con vaina de mielina presentan una conducción saltatoria. Esto se produce porque la vaina se comporta como un excelente aislante que reduce el flujo de iones a través de la membrana en unas 5. 000 veces. Los iones sólo pueden fluir a través de los nódulos de Ranvier y por tanto los potenciales se pueden producir sólo en los nódulos. La conducción saltatoria tiene valor por dos razones: al hacer que la despolarización se conduzca entre nódulos el potencial de acción se hace más rápido, entre 5 y 50 veces. En segundo lugar, ahorra energía para la neurona, porque sólo se despolarizan los nódulos.

VELOCIDAD DE CONDUCCIÓN La velocidad de conducción depende principalmente de dos factores: • La presencia de vaina de mielina hace más rápido el potencial de acción. Las fibras amielínicas conducen a 0, 25 m/seg y las mielínicas hasta 100 m/seg. • El diámetro del axón, a mayor diámetro mayor es la velocidad de conducción.

POTENCIAL EN REPOSO La membrana de cualquier célula presenta una distribución asimétrica de sus iones, así es como en el medio extracelular existe un predominio de Na+ y en el medio intracelular, predomina el K+. Esta diferencia de concentración es mantenida por la bomba Na+ - Ka+ ATPasa con gasto de ATP. Si describimos la situación de otros iones nos encontramos con que hay cloruro (Cl-) en el espacio extracelular, mientras que en la cara interna de la membrana plasmática se acumulan proteínas, sulfatos y fosfatos que le dan un carácter negativo al interior de la membrana neuronal.

GENERACION DEL POTENCIAL DE ACCIÓN FASE 1 POTENCIAL EN REPOSO Este primer paso supone un estado basal en el que aún no se han producido alteraciones que conduzcan al potencial de acción. Se trata de un momento en el que la membrana está a -70 m. V, su carga eléctrica de base. Durante este momento pueden llegar a la membrana algunas pequeñas despolarizaciones y variaciones eléctricas, pero no son suficientes para desencadenar el potencial de acción.
, la estimulación")
FASE 2 DESPOLARIZACIÓN Esta segunda fase (o primera del potencial en sí), la estimulación genera que se produzca en la membrana de la neurona un cambio eléctrico de suficiente intensidad excitatoria (que debe como mínimo generar un cambio hasta los -65 m. V y en algunas neuronas hasta de -40 m. V) como para generar que los canales de sodio del cono del axón se abran, de tal manera que los iones de sodio (cargados positivamente) entran de forma masiva. Las bombas de sodio/potasio (que normalmente mantienen el interior de la célula estable expulsando intercambiando tres iones de sodio por dos de potasio de tal manera que se expulsan más iones positivos de los que entran) dejan de funcionar. Ello generará un cambio de la carga de la membrana, de tal manera que llegue a los 30 m. V. Dicho cambio es lo que se conoce como despolarización.

FASE 2 DESPOLARIZACIÓN Empiezan a abrirse los canales de potasio de la membrana, que al ser también un ión positivo y estar entrando estos masivamente serán repelidos y empezarán a salir de la célula. Esto generará que se frene la despolarización, al perderse iones positivos. Es por ello que como mucho la carga eléctrica será de 40 m. V. Los canales de sodio pasan a cerrarse, y se inactivarán durante un corto periodo de tiempo (lo que impide despolarizaciones sumativas). Se ha generado una onda que no puede volver atrás.

FASE 3 REPOLARIZACIÓN Al haberse cerrado los canales de sodio, este deja de poder entrar a la neurona, a la par que el hecho de que los canales de potasio sigan abiertos genera que este siga siendo expulsado. Es por ello que el potencial y la membrana se hacen cada vez más negativo.

FASE 4 HIPERPOLARIZACIÓN Según sale más y más potasio, la carga eléctrica de la membrana se va haciendo cada vez más negativo hasta el punto de hiperpolarizarse: llegan a un nivel de carga negativa que incluso supera la de reposo. En este momento se cierran los canales de potasio, y vuelven a activarse (sin abrirse) los de sodio. Ello genera que la carga eléctrica deje de bajar y que técnicamente pudiera haber un nuevo potencial, más sin embargo el hecho de que sufre una hiperpolarización hace que la cantidad de carga que sería necesaria para un potencial de acción sea mucho mayor de la habitual. También se reactiva la bomba de sodio/potasio.

FASE 5 POTENCIAL EN REPOSO • La reactivación de la bomba de sodio/potasio genera que poco a poco vaya entrando carga positiva dentro de la célula, algo que finalmente va a generar que vuelva a su estado basal, el potencial de reposo (-70 m. V).

EL POTENCIAL DE ACCIÓN Y LIBERACIÓN DE NEUROTRANSMISORES Este complejo proceso bioeléctrico va a ir produciéndose a partir del cono axónico hasta el final del axón, de tal manera que la señal eléctrica va a ir avanzando hasta los botones terminales. Dichos botones tienen canales de calcio que se abren cuando el potencial llegan a ellos, algo que provoca que las vesículas que contienen neurotransmisores emitan su contenido y lo expulsen al espacio sináptico. Así, es el potencial de acción lo que genera que se liberen los neurotransmisores, siendo la fuente principal de transmisión de la información nerviosa en nuestro organismo.

Fase 1: Reposo Fase 2: despolarización Fase 3: repolarización Fase 4: Hiperpolarización Vuelve a estado de reposo PRE: Periodo refractario
. Psicobiología. Manual")
BIBLIOGRAFÍA • Gómez, M. ; Espejo-Saavedra, J. M. ; Taravillo, B. (2012). Psicobiología. Manual CEDE de Preparación PIR, 12. CEDE: Madrid • Guyton, C. A. & Hall, J. E. (2012) Tratado de Fisiología médica. 12 a edición. Mc. Graw Hill. • Kandel, E. R. ; Schwartz, J. H. & Jessell, T. M. (2001). Principios de neurociencia. Cuarta edición. Mc. Graw-Hill Interamericana. Madrid.
- Slides: 24