Porcentagem de Hidratos de Carbono nos tipos de

Porcentagem de Hidratos de Carbono nos tipos de alimentos mais comuns

Embora a nutrição de nossas células seja oferecida interna e diretamente pelo sangue cujo fluxo é rapidamente regulado, o conteúdo de nutrientes no sangue é afetado também rapidamente pela função diferenciada das células e dos tecidos biológicos. Entre eles o sistema digestório e os alimentos ingeridos e digeridos total ou parcialmente. Nossos costumes foram selecionados (grosseiramente falando) por processos de aprender a buscar e repetir coisas agradáveis e fugir de coisas desagradáveis. Para ambas atividades, nossos antepassados contavam com a necessidade de gastar uma enorme quantidade de energia química de suas reservas (Glicogênio, Lipídeos e mesmo massa muscular de proteínas). Desta forma, a recompensa para este gasto de energia, tinha que ser realmente agradável. Hoje em dia, ela continua a ser crescentemente agradável (resultado de evolução neste sentido, de preparo de alimentos e de imagens publicitárias) mas, o esforço para obtê-las foi enormemente reduzido. Olhando a slide anterior, podemos imaginar que lutar contra este processo básico de luta pelo prazer pode facilmente se transformar numa batalha perdida pois, significa colocar a natureza humana em oposição aos processos mais básicos de sua origem. O correto parece ser aprender a apreciar e desfrutar o prazer com moderação, com educação e cuidadosamente e, aumentar a atividade física diária. Deslocar o padrão da dieta na direção dos alimentos que aportam carbo-hidratos de fácil digestão e absorção (a absorção deverá ser sempre rápida para impedir a perda de água por desidratação - osmose) para aqueles mais difíceis sem eliminar por completo aqueles mais apreciados. E cuidado especial para com aqueles como o açúcar no cafezinho que se mostra no dia a dia imperceptível.

Reações da via glicolítica com destaque para fornecimento de energia química na forma de ATP e de poder redutor. A conversão de piruvato em lactato mantém ativo o sistema de transferência de poder redutor de forma independente do ciclo de Krebs e da cadeia respiratória encontradas na mitocôndrias

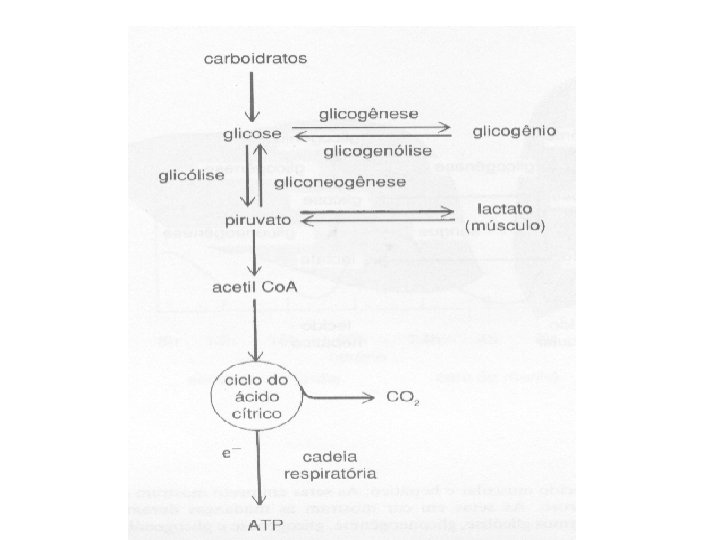
Ciclo de Krebs – Aerobiose Na presença de mitocôndrias e oxigênio os carbonos do piruvato fornecem o Acetil do Acetil Co. A e podem fornecer ATP a partir da cadeia respiratória ligada ao ciclo de Krebs Acetil Co. A OXALOACETATO CITRATO

Ciclo de Krebs e Cadeia Respiratória • Para a bioquímica não se trata de uma via metabólica e sim de um ciclo pelo fato de que o ponto inicial e o ponto final são idênticos: o mesmo oxaloacetato. • Entretanto, trata-se de um sistema de enorme importância para todas as vias de metabolismo aeróbico que possam fornecer Acetil Co. A para serem oxidados e produzirem ATP na cadeia respiratória usando a energia acumulada no potencial de membrana mitocondrial. • Este ciclo também permite que a glicose forneça carbonos para a produção de amino acidos não essenciais e em condições de dieta hiper proteica permite que os carbonos dos amino ácidos glicogênicos sejam convertidos em glicose. • A cadeia respiratória, também não é uma via metabólica mas, será o sistema responsável pelo processo mais eficiente de conversão de energia de oxido-redução em síntese de ATP repare que transferências de poder redutor (NADH e FADH 2) ocorrem depois da produção de gás carbônico a energia é extraída da transferência de elétrons e não da quebra de ligações carbono-carbono. . • Ciclo de Krebs e cadeia respiratória operarão em conjunto para manter os níveis intracelulares de ATP com velocidade de queima de moléculas de acetil Co. A em baixa velocidade. Ele é controlado pela oferta de ADP (ATP gasto pela célula)

Cada NADH possui energia livre para obter 3 moléculas de ATP. Já o FADH 2 produz 2 de ATP.

VIA GLICOLÍTICA SEM REQUERIMENTO DE OXIGÊNIO Citrato torna enzima 4 inibível por ATP RENDE 2 ATP POR MOLÉCULA DE GLICOSE CONVERTIDA EM 2 DE LACTATO CITRATO ALTO INDICA GRANDE PRODUÇÃO DE ATP NO CICLO DE KREBS E CADEIA RESPIRATÓRIA DENTRO DA CÉLULA.

Embora consideremos que todos os carbonos dos componentes de uma célula viva estejam em contato com o ambiente extracelular e portanto, participem da abertura termodinâmica oferecida pela nutrição, os fluxos de carbono do que consideramos metabolismo não são idênticos. Assim, os carbonos da glicose podem entrar e sair das células vivas em grande velocidade na forma de CO 2 ou de lactato o mesmo não ocorrerá com os carbonos dos amino ácidos que compões as proteínas que serão nossas enzimas ou mesmo com outras moléculas orgânicas requeridas para as transformações do metabolismo como são as co-enzimas ou co-fatores como NAD, NADP, FAD e/ou Co. A. No caso dos transportadores de H (NAD, NADP, FAD), eles acoplarão direta ou indiretamente (quando cruzamento de barreiras/membranas são necessários) reações de oxido redução. Apenas os H doados ou recebidos ocorrerão com alterações químicas, em alta velocidade. Os carbonos de NAD, NADP etc. , serão trocados à velocidade muito baixa. No caso, da Co. A, o acetato ou succinato que poderão ser ativados pelo Co. A também participarão de fluxos rápidos enquanto que os carbonos da molécula de Co. A serão em sua grande maioria mantidos por longo tempo ou seja participarão de fluxos muito lentos. Por esta razão eram denominados de co-enzimas pois, apresentavam a mesma capacidade que as enzimas apresentam de terem a maior parte de suas estruturas preservadas durante um número enorme de reações catalisadas. E apenas não eram considerados partes fixas das enzimas por não estarem unidas de forma contínua a elas por ligações covalentes como as que unem um amino ácido ao outro (ligação peptídica). Assim a notação que melhor descreve os transferidores de H é coloca-los abaixo da seta que representaria uma enzima como um par de oxido-redução que obedece no esquema a forma que recebe o H junto ao substrato que o doa e a forma que doará o H junto ao produto que o perdeu. O ATP, ADP e AMP mereceriam a mesmas considerações mas, com a ressalva de que é objeto de uma regulação específica sem relação estrita com uma única etapa do fluxo. Esta regulação observa ao longo da evolução é denominada “Carga energética” como se fosse a carga de uma “bateria química” participante da maioria dos processos onde variação de energia livre pode ser obtida ou requerida.

Efeito do Etanol sobre NADH + H

Compartimentos Extracelular Intramitocondrial

Mesmo com este esquema bem simplificado para os compartimentos existentes dentro da célula, fica clara a maior complexidade em relação à bioquímica da bactérias. Se a bioquímica de outros microrganismos mais complexos como fungos (eucariontes) forem usados para comparação, ainda assim, é importante destacar que as células destes eucariontes não regulam aspectos físico-químicos de seu meio extracelular como ocorre com nossas células em relação a por exemplo, homeostasia glicídica (manutenção dos níveis de glicose no sangue). Mesmo com esta apresentação simplificada de nossos compartimentos intracelulares é possível entender a importância da separação entre o meio intramitochondrial e extramitocondrial em nosso citoplasma. Nossas mitocôndrias transformam em potencial energético as oxido-reduções do metabolismo e usam este potencial de forma controlada para obtenção de ATP. Desta forma, não queimamos todas as moléculas orgânicas que obtemos por nossa nutrição. Queimamos apenas na medida em que necessitamos fazer ATP a partir de ADP que aparece quando ATP gasto. Foi este mecanismo de controle respiratório que nos possibilitou fazer as reservas que usamos para as atividades que nos agradaram durante a evolução de nossos ancestrais. Atualmente, os fatores que nos motivaram a executar trabalho por um mecanismo de memória que os associa com a recompensa agradável, são mais fortes e o trabalho requerido para sua obtenção são incrivelmente menores. O mesmo mecanismo de obtenção de reserva que salvou nossos antepassados, nos dá facilmente, a obesidade epidêmica.

Preparação mitocondrial de fígado de ratos - Registro analógico original – Clark (Deptos de Bioquímica e de Cirurgia Lab Cirurgia-experimental FMRP-USP-1986) Mesmo com oferta de substrato, de oxigênio, de enzimas e de genoma presentes, a respiração (consumo de oxigênio e produção de gás carbônico) só se ativa com ADP. Quando todo o ADP foi transformado em ATP se inibe de novo (estado IV). A queima de substratos (HC, lipideos, aminoàcidos) mitocondrial é ligada à queima ou necessidade de ATP.

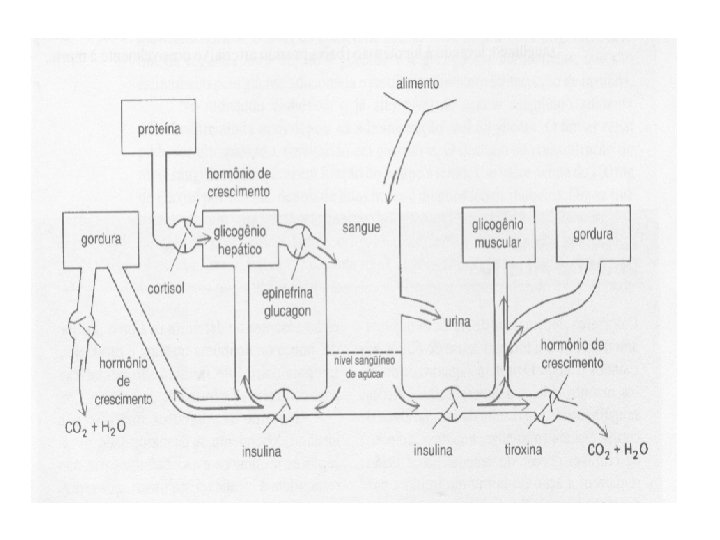
Reservas de hidrato de carbono Glicogênio muscular não é afetado pelo jejum e sim pelo exercício (reserva de ATP) Glicogênio hepático acaba em poucas horas de jejum (reserva de glicose)

Veja como glicogênio hepático vai sendo lentamente alterado por períodos dentro do dia de jejum e por absorção de hidratos de carbono provenientes dos alimentos

Na madrugada, já sem glicogênio hepático é a via neoglicogenética que mantém os níveis de glicose circulante. Nas dietas sem hidratos de carbono (hiperproteicas) esta via serve para fabricar glicogênio ou açúcar do leite ou ainda, manter a glicemia. Dentro de uma célula, quando esta via estiver ativada, a via glicolítica estará inibida para impedir quebra fútil de ATP. Neoglicogênese

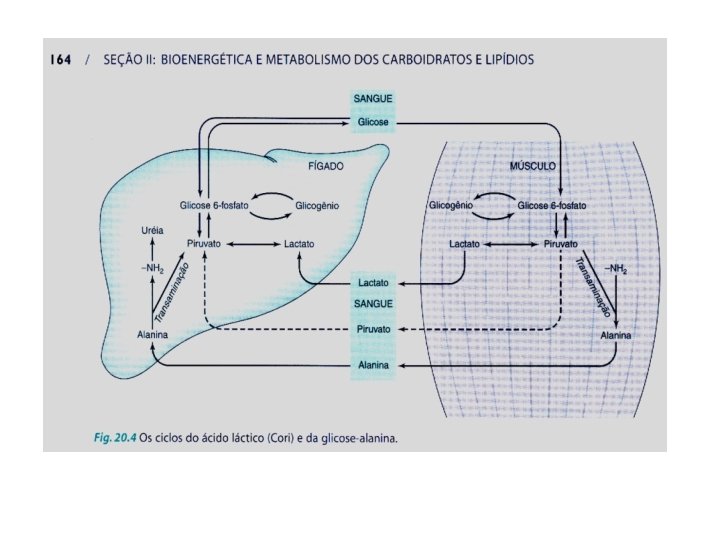
Integração metabólica entre órgãos. A regra que numa célula, impede o funcionamento simultâneo das vias neoglicogenética e glicolítica não impede o funcionamento simultâneo das duas vias em órgãos diferentes. No primeiro caso, observa-se regulação de ATP (intracelular) e no segundo, regulação da glicemia (extracelular). Glicogênio muscular reserva de ATP Mantém ATP dentro das células (intracelular) Glicogênio hepático reserva de glicose Mantém a glicose circulante – glicemia (extracelular)
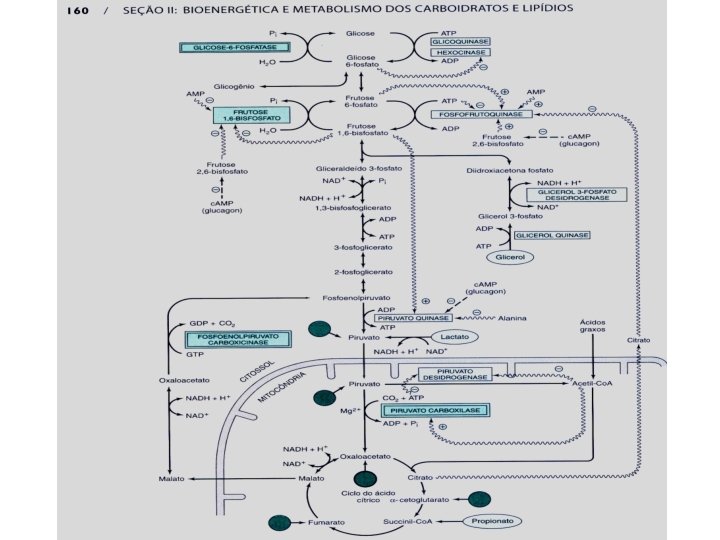

Inibição da via glicolítica por ATP + Citrato Na projeção anterior, aparece uma seta que indica que o Citrato (a primeira molécula formada no Ciclo de Krebs) inibe a via glicolítica. Na realidade, ele torna a enzima regulatória (4) da via glicolítica inibível por concentrações normais de ATP ou seja, desde que existam outras fontes de Acetil Co. A para o ciclo de Krebs e a produção de ATP seja mantida a via glicolítica terá sua velocidade reduzida. Isto significaria que os ATPs formados por lipídeos (Acidos graxos livres – AGL) ou por aminoácidos cetogênicos são melhores do que o ATP formado a partir do piruvato proveniente da glicose? Não! Mostra de outra forma o que já havíamos percebido na regulação de músculo e fígado que a regulação do intracelular é uma (Mantêm ATP) e a do extracelular (manutenção da glicemia) é outra. A própria célula não recebe benefício por fontes alternativas de ATP mas, as células como os glóbulos vermelhos (hemácias) e os que dependem do metabolismo da glicose são beneficiadas pela homeostasia (sentido correto).



Considerações finais • Abordamos dois aspectos do metabolismo: No primeiro aspecto, observamos o fluxo dos átomos de carbono de uma molécula como o CO 2 para outra molécula como a de glicose na fotosíntese. Ou alternativamente, o fluxo dos carbonos de glicose para o CO 2 do ar pela via glicolítica aeróbica. No segundo aspecto, observamos que o metabolismo não é dado apenas pelas transferências de átomos de carbono de uma molécula que inicia uma via metabólica para a molécula que é o produto final desta via. Deve-se considerar adicionalmente, os processos de fornecimento de energia seja na forma de síntese de ATP, de potencial de óxido-redução ou de liberação de calor. Finalmente, percebemos como estes aspectos se inter-relacionam quando se observa o efeito do metabolismo do etanol. O etanol pode ser ingerido como fonte rápida de calor. Mas, afeta uma via metabólica biosintética, a neoglicogênese, devido a alteração de oxido-redução provocada pelo metabolismo do etanol. Este último aspecto, fica bem importante quando consideramos que em madrugada fria (sem ingestão de alimentos acompanhando) uma pequena ingestão de álcool “para aquecer” pode causar súbita hipoglicemia ao interferir na neoglicogênese hepática no momento, em que esta via se torna a principal reguladora da glicemia. Além da noção de espaço e compartimentos celulares a noção de TEMPO é fundamental em bioquímica em geral e não apenas para o funcionamento do cérebro. As regulações rápidas mantem a vida e ocorrem sem depender de alterações da expressão gênica ( leituras de DNA). .
- Slides: 25