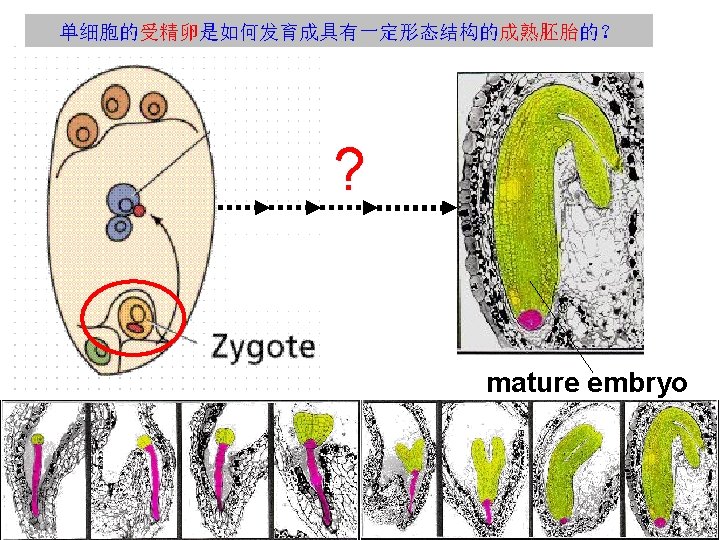
Plant Embryogenesis Pattern Formation from a Single Cell

Plant Embryogenesis 植物的胚胎发生 Pattern Formation from a Single Cell 蒋家敏 200800140063
 4. 表皮原期(Dermatogen stage) 5. 球形胚期(Globular stage)")
胚胎发育的主要阶段 1. 受精卵的不均等分裂 2. 2细胞期 3. 8细胞期(Octant stage) 4. 表皮原期(Dermatogen stage) 5. 球形胚期(Globular stage) 6. 早心形期(early heart stage) 7. 心形胚期(Heart stage) 8. 鱼雷胚期(Torpedo stage) 9. 手杖胚期(Bent cotyledon stage)



 At the transition from the octant stage embryo, a single round")
4. 胚胎的表皮原期(Dermatogen stage) At the transition from the octant stage embryo, a single round of tangential divisions(切向分裂) separates an outer layer of eight epidermal precursor (or protoderm) cells from eight inner cells.
 Protoderm and inner cells soon become histologically distinguishable and due to")
5. 球形胚期(globular stage) Protoderm and inner cells soon become histologically distinguishable and due to predominantly anticlinal divisions(垂周分裂) in the outer layer, the protoderm remains essentially separated from inner cells throughout development.
 At late globular stage, when the number of cells has")
6. 早心形期(early heart stage) At late globular stage, when the number of cells has increased to more than a hundred, the embryo gradually assumes a triangular shape due to localized growth at two opposite positions in the apical region. The early heart stage embryo (also referred to as ‘triangular’ stage) comprises approximately 200 cells and the primordia of most major seedling organs, cotyledons, hypocotyl and primary root, as well as the basic tissue types, provascular, protoderm and cortex are anatomically discernible.
 此时胚胎呈心形,子叶从胚胎两侧长出。随后,长出的子叶之间的细胞开始形 成初级的茎分生组织。 In post heart-stage embryos the shoot meristem becomes")
7. 心形胚期( heart stage) 此时胚胎呈心形,子叶从胚胎两侧长出。随后,长出的子叶之间的细胞开始形 成初级的茎分生组织。 In post heart-stage embryos the shoot meristem becomes discernible as three distinct cell layers that will subsequently attain a tunica-corpus (原套-原 体)organization (Barton and Poethig, 1993).
及手杖胚期(Bent-cotyledon stage) In torpedo and bent-cotyledon stage embryos, provascular tissues(原形成层组 织) also")
8. 鱼雷期(Torpedo stage)及手杖胚期(Bent-cotyledon stage) In torpedo and bent-cotyledon stage embryos, provascular tissues(原形成层组 织) also become recognizable within cotyledon primordia and the cellular organization of hypocotyl(胚轴) and root is completed. Although cells in most tissues will complete differentiation after germination, the complexity of the tissue pattern in the bent-cotyledon stage embryo basically equals that of the seedling.
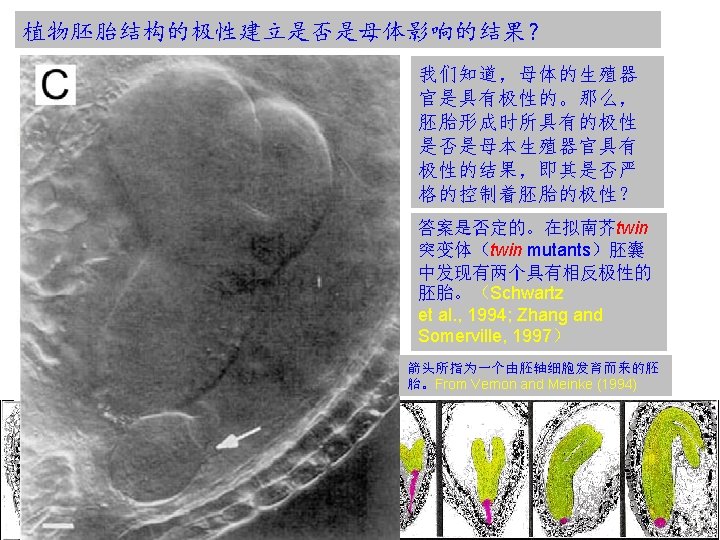

植物胚胎极性是细胞谱系还是位置决定的结果? The reproducible sequence of cell divisions in the Arabidopsis embryo effectively results in the specification of cell fates along predictable cell lineages. For example, characteristic early divisions suggest a hierarchy of partitioning events, in which early established lineages contribute only to particular structures in the mature embryo. Does this predictability reflect lineage-imposed cell fate restriction? However, in Arabidopsis mutants where the regular sequence of cell divisions is dramatically disturbed and major organs and tissues remain properly positioned. These, and many similar observations suggest that the specification of the basic embryonic pattern is largely, if not entirely, based on positional cues( 位置线索), which are remarkably indifferent to cell boundaries, cell numbers and overall dimensions. However, it does not exclude a role for lineage-dependent cell fate specification in local patterning processes at later stages.
的形成 The main elements to be positioned along the apical-basal axis are the")
顶-基向模式(Apical-basal Pattern)的形成 The main elements to be positioned along the apical-basal axis are the shoot apical meristem (SAM)(茎尖分生组织), cotyledons(子叶), hypocotyl(胚轴), radicle(幼根) and the root apical meristem (RAM)(根尖分生组织). SAM和部分子叶来源于u. t(upper tier);胚 轴,幼根来源(包括大多数的RAM起始)于 l. t(lower tier).
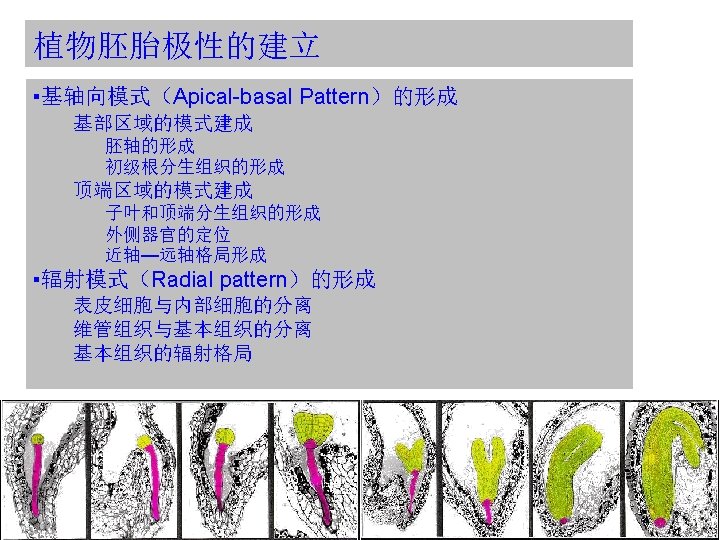
的形成 Apical-basal polarity Patterning the basal domain Formation of the embryo axis Formation")
基轴向模式(Apical-basal Pattern)的形成 Apical-basal polarity Patterning the basal domain Formation of the embryo axis Formation of the primary root meristem Patterning the apical domain Generating the cotyledons and the shoot apical meristem Positioning of lateral organs Adaxial-abaxial patterning
 WOX: WUSCHEL RELATED HOMEOBOX WUSCHEL(WUS) : Homeobox gene controlling the stem cell")
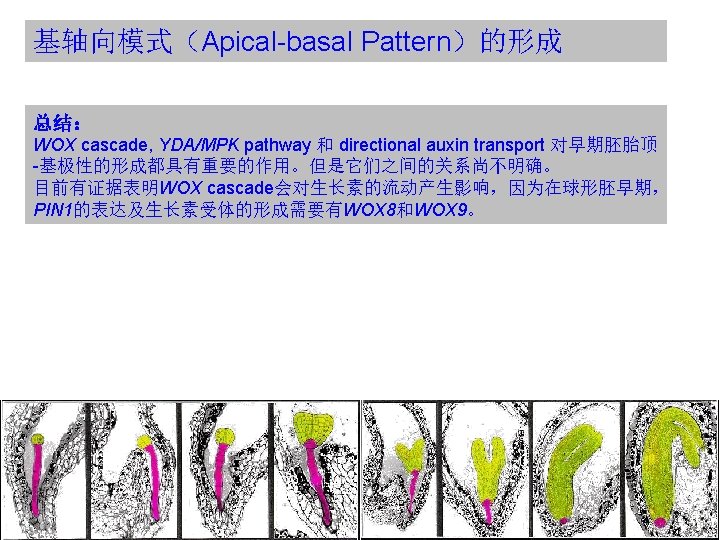
WOX cascade(WOX级联) WOX: WUSCHEL RELATED HOMEOBOX WUSCHEL(WUS) : Homeobox gene controlling the stem cell pool. Expressed in the stem cell organizing center of meristems. Required to keep the stem cells in an undifferentiated state. Regulation of WUS transcription is a central checkpoint in stem cell control. Homeobox(同源框基因): The homeobox is a stretch of DNA about 180 nucleotides long that encodes a homeodomain. Homeodomain(同源异型结构域): A domain in a protein that is encoded for by a homeobox, that consists of about 60 amino acid residues which are usually similar from one such domain to another, and that recognizes and binds to specific DNA sequences in genes regulated by the homeotic gene.
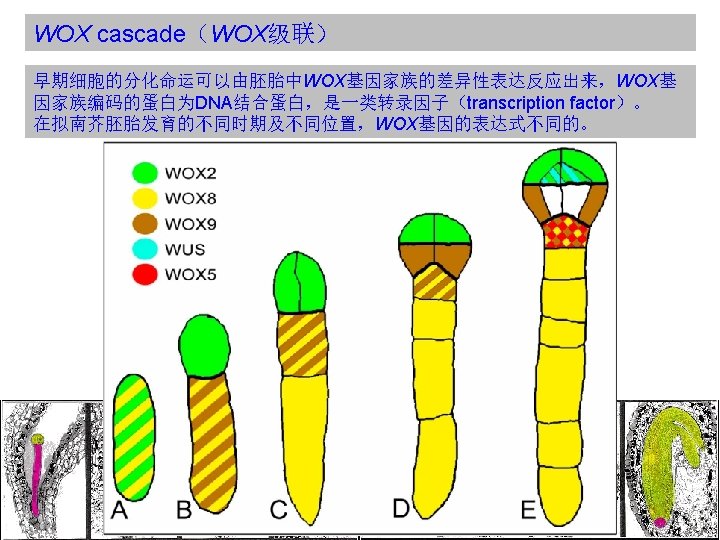
 Functions of WOX 8 and WOX 9 in the apical domain appear")
WOX cascade(WOX级联) Functions of WOX 8 and WOX 9 in the apical domain appear to be at least partly mediated by downstream activation of WOX 2 expression, which is absent in the embryo of wox 8 wox 9 double mutants. WOX 2 expression alone appears sufficient to confer important characteristics of the apical cell lineage. Misexpression of WOX 2 in the wox 8 wox 9 background reduces the asymmetry of the initial division of the zygote and promotes aspects of the apical lineage in the basal cell. In wox 8 wox 9 double mutants, WOX 2 and other apical cell lineage features are not expressed. Expression of WOX 2 seems to be instrumental in apical fate acquisition, because targeted mis-expression of WOX 2 confers apical-cell features.
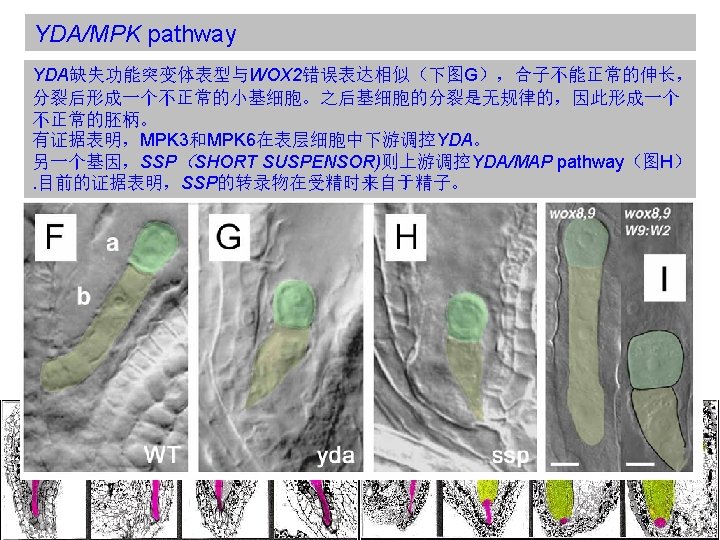

YDA/MPK pathway Mutant analysis suggests that WOX-dependent regulation and the MPK/YDA genes do not operate on a single linear pathway. (Breuninger et al. , 2008) 虽然WOX类突变体和 YDA/MPK类突变体的表型很 类似,但是它们是通过不同 的信号途径起作用的。
 PIN:PINFORMED family of auxin efflux facilitator In the absence of more")
Directional auxin transport(生长素的方向性运输) PIN:PINFORMED family of auxin efflux facilitator In the absence of more direct evidence for the position and polarity of major auxin transport routes, such polar avenues decorated by PIN protein expression have been used to establish plausible models linking dynamic patterns of auxin transport to patterning processes in all parts of the plant, beginning with the formation of the main body axis in the early globular embryo. 胚胎中的PIN蛋白可以使用合成的生长素响应启动子元件DR-5显示。
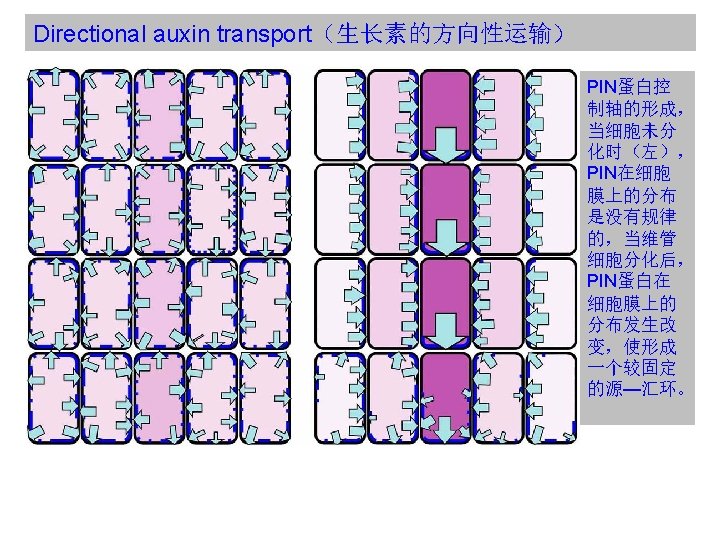
 In the early stages of embryogenesis, localization of the PIN 7")
Directional auxin transport(生长素的方向性运输) In the early stages of embryogenesis, localization of the PIN 7 auxin effl ux facilitator (cyan) to the apical membranes of basal cell and subsequently suspensor cells appears to drive auxin fl ux (arrows) upwards. A weak DR-5 marked auxin perception maximum (light purple) suggestive of auxin accumulation is seen in the apical parts of the developing embryo up to and including the dermatogen stage. During the globular stage PIN 1 (dark blue) localization in the basal membranes of the inner cells of the embryo is associated with a switch in the apparent direction of auxin fl ux to apical – basal. At this time PIN 7 is now seen concentrated in the basal membranes of the hypophysis and suspensor cells. A stronger auxin perception maximum (purple) also appears in the hypophysis and apical cells of the suspensor, which becomes restricted to the daughter cells of the hypophysis. PIN 1 distribution in the cells of the L 1 layer is associated with the generation of auxin perception maxima at the positions of incipient cotyledon initiation.
 Further evidence supporting the critical role played by directional auxin transport")
Directional auxin transport(生长素的方向性运输) Further evidence supporting the critical role played by directional auxin transport in establishing apical-basal polarity is provided by work with the EMBRYO DEFECTIVE 30/GNOM (EMB 30/GN) mutant. EMB 30/GN突变体幼苗呈球状,细胞完全分化,但是分裂方向是随机的,维管细胞 仍处于中心,说明其为选择性地丢失顶-基极性。 EMB 30/GN encodes an Adenosyl ribosylation factor(腺苷核糖基化因子) Guanine nucleotide Exchange Factor(鸟嘌呤核苷酸交换因子)(ARF GEF), which functions as an endosomal regulator of vesicle budding. 为什么EMB 30/GN调控囊泡的出芽(vesicle budding)会影响生长素的运输? PIN复合体的循环依赖于肌动蛋白驱动的质膜和内膜区室间的囊泡运输。
的形成 Apical-basal polarity Patterning the basal domain Formation of the embryo axis Formation")
基轴向模式(Apical-basal Pattern)的形成 Apical-basal polarity Patterning the basal domain Formation of the embryo axis Formation of the primary root meristem Patterning the apical domain Generating the cotyledons and the shoot apical meristem Positioning of lateral organs Adaxial-abaxial patterning

Formation of the embryo axis The initiation of the embryo axis occurs through strictly oriented divisions in the early globular embryo. Divisions oriented along the apical-basal axis predominate among l. t. cells and generate a cylinder of parallel cell files.

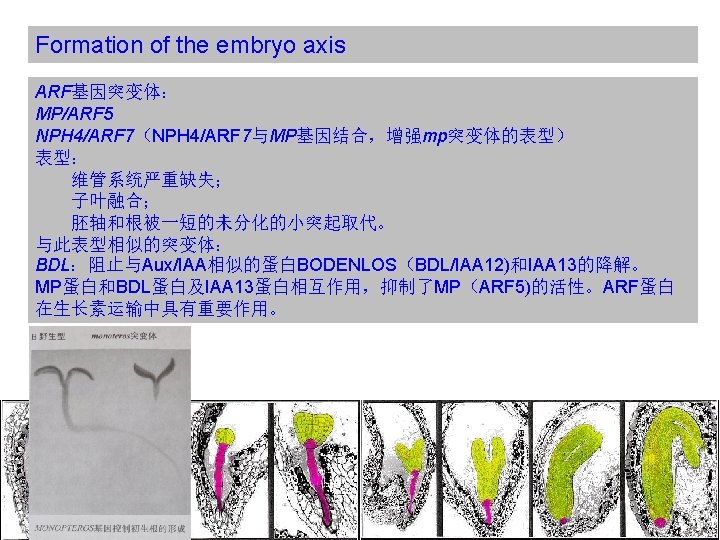
Formation of the embryo axis Auxin signaling and directional auxin transport are essential for normal patterning in the basal domain. ARF: AUXIN RESPONSE FACTOR Aux/IAA: Auxin/Indole Acetic Acid TIR 1: TRANSPORT INHIBITOR RESPONSE 1
突变体与mp和bdl突变体相反,它们没有顶端部分,半显性的tpl突变体 可以抑制bdl突变表型。TPL也已经被证明可以通过一个乙烯响应因子(ERF)与 BDL/IAA 12作用,从而调控MP/ARF 5蛋白的活性。 A, E, I: 野生型")
Formation of the embryo axis tpl(topless)突变体与mp和bdl突变体相反,它们没有顶端部分,半显性的tpl突变体 可以抑制bdl突变表型。TPL也已经被证明可以通过一个乙烯响应因子(ERF)与 BDL/IAA 12作用,从而调控MP/ARF 5蛋白的活性。 A, E, I: 野生型 B, F, J: tpl-1突变体 C, G, K: bdl突变体 D, H, L: tpl-1, bdl双突变体 Overall therefore, auxin signal transduction and regulated auxin transport are required for oriented cell divisions and vascular differentiation in the basal domain, and these patterning events constitute prerequisites for subsequent radial differentiation and root meristem formation.

Formation of the primary root meristem 位置信号途径:It has long been suspected that cell fate in the RAM is largely controlled by positional cues and single-cell ablation has been employed to trace the origin of some of these cues。 One, originating from the QC, seems to confer stem cell identity to cells surrounding the QC. Another type of positional signal seems to help integrate the tissue identity of all cells in a given layer. Blue arrows indicate that the acquisition of QC cell fate seems to be dependent on signals from the shoot, red arrows the dependence of stem cell fate on signals from the QC. Black arrows represent endodermis inducing signals from the stele (Figure 18) and green arrows the stabilization of tissue identity within each layer.

Formation of the primary root meristem 如果静止中心决定了其周围细胞的命运,那么QC自己又是如何定位的? Auxin supplied from the proembryo and polarly transported in an apical basal direction results in the formation of an auxin response maxima in the nearby suspensor cells, the uppermost of which assumes a hypophyseal fate. The lensshaped daughter cell of the hypophysis subsequently gives rise to the QC.

Formation of the primary root meristem Another intriguing network of genes involved in the establishment of the RAM is the PLETHORA (PLT) gene family.
 and SCARECROW (SCR) also")
Formation of the primary root meristem The GRAS-domain genes SHORTROOT(SHR) and SCARECROW (SCR) also play a critical role in the specifi cation of the QC fate.

Formation of the primary root meristem Another identified factor in the QC establishment and maintenance is WOX 5. 此外,细胞分裂素与生长素相互拮抗,在胚胎模式建成中具有重要作用。
的形成 Apical-basal polarity Patterning the basal domain Formation of the embryo axis Formation")
基轴向模式(Apical-basal Pattern)的形成 Apical-basal polarity Patterning the basal domain Formation of the embryo axis Formation of the primary root meristem Patterning the apical domain Generating the cotyledons and the shoot apical meristem Positioning of lateral organs Adaxial-abaxial patterning
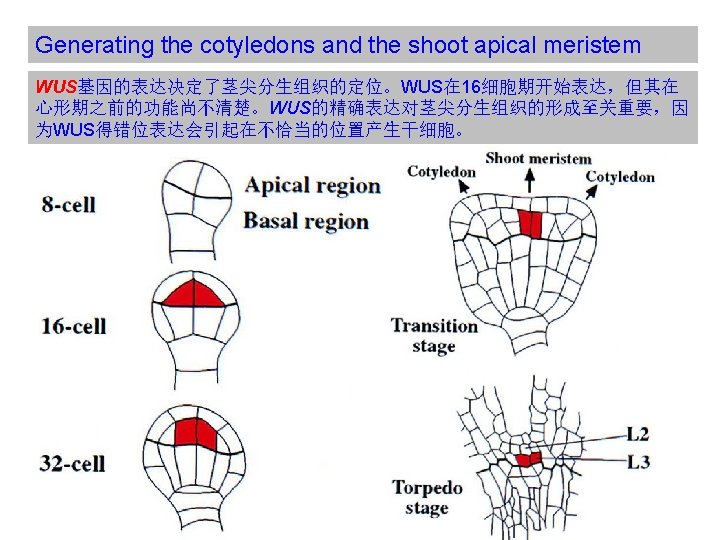

Generating the cotyledons and the shoot apical meristem WUS基因的表到受到CLAVATA pathway的负调控。CLAVATA由三个基因构成, CLV 1编码一个受体激酶,CLV 2则编码一个类似受体激酶的蛋白,与CLV 1形成异 二聚体,CLV 3则与CLV 1/CLV 2复合体结合,为CLV 1的受体。 (A) Cells in the outer layer of the meristem secrete CLV 3 protein, which binds to CLV 1 receptor proteins on target cells in an adjacent, more central region of the meristem, presumably stimulating the differentiation of the target cells. (B) Some parts of the intracellular signaling pathway activated by CLV 3 binding. The CLV receptor protein is thought to be a homodimer or heterodimer, which phosphorylates itself on serines and threonines, thereby activating the receptor and leading to the activation of a Rho-like GTPase. The signaling pathway after this point is unclear, but it leads to the inhibition of a gene regulatory protein in the nucleus, thereby blocking the transcription of genes that otherwise might inhibit differentiation. The phosphatase dephosphorylates the receptor and thereby negatively regulates the signaling pathway. (From Signaling in Plants)
 In situ hybridizations of STM")
Generating the cotyledons and the shoot apical meristem (A–F) In situ hybridizations of STM (A, B), CUC 2 (C, D) and AS 1 (E, F), in globular (A, C, E) and heart (B, D, F) stage embryos. Arrowheads in C and D point to the protoderm, where CUC 2 is absent. The arrowhead in F indicates the SAM initials (H) Model of STM/CUC/AS interactions. The expression domains of STM and CUC 2 (and the other CUC genes) are largely overlapping at the globular stage. Activity of CUCs promotes STM expression (A, C). Conversely, STM downregulates the CUCs. (B, D). STM activity promotes the establishment and maintenance of the SAM. This is in part accomplished by another function of STM: to inhibit expression of the genes AS 1 and AS 2 (here represented by AS 1) in the SAM. The counter-acting activities of CUCs and STM lead to the formation of a torus of CUC expression around a STM domain centered on the SAM in the bent cotyledon stage, depicted in I. (I) Schematic transverse section through the apex of a bent cotyledon embryo showing the inner STM expression domain surrounded by the CUC 2 domain.

Generating the cotyledons and the shoot apical meristem There are several other genes whose expression is important for proper SAM and cotyledons formation. GURKE (GRK):is an acetyl-Co. A carboxylase(乙酰辅酶A羧化酶), suggesting metabolites derived from malonyl-Co. A(丙二酰辅酶A) could be important for patterning in apical domain. PASTICCINO (PAS):is a FK 506 -binding protein: is a family of proteins that have prolylisomerase(脯氨酸 异构酶) activity and are related to the cyclophilins(亲环素蛋白) in function, though not in amino acid sequence. function as protein folding chaperones for proteins containing proline residues. 证据表明,PAS与NAC-like转录因子FAN相互作用,使其能够进入细胞核阻抑细胞 分裂素依赖的细胞分裂。

Pisitioning of lateral organs Several lines of evidence support a central role for auxin in shoot lateral organ initiation. First, mutations in auxin signal transduction genes MP, BDL, AUXIN RESISTANT 6/CULLIN 1 (AXR 6/CUL 1) result in cotyledon fusions. loss of both MP/ARF 5 and NPH 4/ARF 7 completely prevents the formation of cotyledon primordia. Second, lateral organ formation does not only require proper auxin perception but also auxin transport. Finally, local auxin application has been shown to induce shoot organ formation at the site of application.

Pisitioning of lateral organs A–D: Model for lateral organ initiation by auxin. F: Model for positioning of cotyledons.

Pisitioning of lateral organs In post-embryonic growth, correct positioning of lateral organ initiation by auxin transport mediated processes has also been shown to depend on a family of auxin influx proteins. AUXIN PERMEASE 1 (AUX 1) LIKE AUX 1, 2 3 (LAX 1, 2, 3) Generally, the influx carriers appear to buffer PIN-mediated patterning.

Pisitioning of lateral organs Mutations affecting auxin synthesis have also been found to impact on lateral shoot organ positioning. There are eleven members of the flavin monooxygenase(黄素单加氧酶) family YUCCA-LIKE(YUC genes) in Arabidopsis, which catalyze a rate limiting step in auxin biosynthesis. 生长素在子叶和外侧器官形成过程中具有十分重要的作用!

Adaxial-abaxial patterning The development of laminar lateral organs involves an additional patterning step, the specification of adaxial(近轴) and abaxial(远轴) cell identities. There are two observations which are important for the discussion of apical-basal patterning in the embryo. First, it has long been recognized that dorso-ventral patterning uses apical-basal polarity as a reference , evidenced by the fact that lateral organs fail to differentiate adaxial-abaxial polarity if insulated from apical signals through vertical incisions in the meristem. Secondly, adaxial-abaxial patterning seems to feed back on the regulation of apical-basal patterning.

Adaxial-abaxial patterning 许多的基因网络涉及到植物胚胎近轴-远轴模式的形成: The TCP expression domain defines the lateral side and is bordered by the CUC expression domain encircling the SAM. The class III HD-ZIP, along with the KANADI (KAN) and YABBY (YAB) genes play acritical role in the process. class III HD-ZIP: Another class of transcription factors Expression of the class III HD-ZIP marks the central domain of the embryo from the globular stage and onward. The KANADI genes, KAN 1, 2, and 3 (but not KAN 4) are initially expressed throughout the early globular stage embryo, but their domain of expression becomes restricted to the basal peripheral region. The YAB genes seem to specify abaxial fate in cotyledon primordia in a highly redundant manner.
 In situ hybridization of REV (A, B), FIL (C, D) and")
Adaxial-abaxial patterning (A–F) In situ hybridization of REV (A, B), FIL (C, D) and KAN 1 (E, F), in globular (A, C, E) and heart (B, D, F) stage embryos. Images A, B reproduced from Emery et al. (2003) with permission from Elsevier Limited; C, D reproduced/adapted from Siegfried et al. (1999) with permission from The Company of Biologists; E, F reproduced with permission from Macmillan Publishers Ltd: Nature, Kerstetter et al. (2001). (G) Schematic representation of the expression domain shown in A-F. The class III HD-ZIP genes, here represented by REVOLUTA, are expressed in the central domain at the globular and heart stage, while the YABBY genes, represented by FILAMENTOUS FLOWER, and the KANADI genes, represented by KANANDI 1, mark the peripheral domain. The activity of the class III HD-ZIPs promotes a central/adaxial fate in their domain of expression, which is also crucial for initiation of the SAM, while the KAN/YAB genes promote a peripheral/abaxial fate simultaneously limiting the expansion of the SAM. The balance of these antagonistic activities is critical for aspects of radial patterning in the embryo axis as well as for the appropriate adaxial/abaxial polarity in lateral organs, including cotyledons.
的形成 The first radial patterning event at the dermatogen-stage separates a surface layer")
辐射模式(Radical Pattern)的形成 The first radial patterning event at the dermatogen-stage separates a surface layer and inner cells of a sphere, while subsequent radial patterning occurs within the radial symmetry of a cylinder. Therefore, it is possible that the separation of the epidermis and subsequent radial patterning are mechanistically unrelated.
的形成 Separating epidermal and inner cells Separating vascular and ground tissue Radial patterning")
辐射模式(Radical Pattern)的形成 Separating epidermal and inner cells Separating vascular and ground tissue Radial patterning in the ground tissue

Separating epidermal and inner cells The mechanism responsible for differential cell fate acquisition by outer and inner cells at the dermatogen stage is not clear, and no mutation has been identified that completely abolishes this first step in radial patterning. Several models can be envisaged: Concentration gradient of an unknown signal molecule could function as a morphogen(形态素) to specify cell fates in the radial dimension. External signal from the endosperm could specify epidermal fate.
 Images of fl uorescent reporter constructs for the")
Separating epidermal and inner cells (C–F) Images of fl uorescent reporter constructs for the expression of At. ML 1. C: At. ML 1 reporter expression in both the apical and basal cell after division of the zygote. (D) At the dermatogen stage the reporter has been downregulated in the inner cells. (E, F) In the late globular and heart stages respectively, the At. ML 1 reporter marks the outer protodermal layer.
 genes, ARABIDOPSIS CRINKLY 4 (ACR")
Separating epidermal and inner cells Two receptor-like kinase (RLK) genes, ARABIDOPSIS CRINKLY 4 (ACR 4) and ABNORMAL LEAF SHAPE 2 (ALE 2) , have also been implicated in the development of surface cells. ACR 4和ALE 2的突变在原表皮分化中具有协同效应(synergistic effect), arc 4 ale 2 双突变体不能保持同时表达PDF 2和At. ML 1. ACR 4和ALE 2蛋白在体内能够相互磷酸化,表明它们在同一条途径中起作用。(? ) The expression of several epidermal genes, including ACR 4, is dependent on At. ML 1 and PDF 2, which may bind the L 1 cisacting element in the promoters of these genes.
: 膜整合蛋白,在 16细胞期帮助维持PDF 2和")
Separating epidermal and inner cells DEFECTIVE KERNEL 1 (At. DEK 1): 膜整合蛋白,在 16细胞期帮助维持PDF 2和 At. ML 1基因的表达域。 Loss of At. DEK 1 eliminates At. ML 1 and ACR 4 expression and prevents the formation of a defined protoderm. RECEPTOR-LIKE PROTEIN KINASE 1 (RPK 1)和TOADSTOOL 2(TOAD 2)基因, 作用不明确,a double mutant rpk 1 toad 2 shows excessive cell division, aberrant protodermal cell morphologies and subsequently, embryonic arrest. RPK 1 has been previously reported as involved in ABA signaling and TOAD 2 is its closest homolog. Other Leucine-Rich Repeat RLK, such as CLV 1, have been shown to be involved in patterning.

Separating vascular and ground tissue Proper patterning of the vasculature relies on functional auxin signaling, as illustrated by arf 5/mp and iaa 12/bdl mutants. Mutations in class III HD-ZIP genes cause defects in the vascular patterning in the shoot and ectopic expression of KAN 1 or KAN 2 prevent the formation of vasculature in the cotyledons and hypocotyl.

Separating vascular and ground tissue Cytokinins have been implicated in cell division and cell differentiation within the vascular cylinder. AHK 4/CRE 1/WOL encodes a membrane associated two-component histidine kinase and has been identifi ed as a cytokinin receptor by conferring cytokinin responsiveness to yeast cells, when inserted into an appropriate yeast signal transduction pathway. WOL突变导致根分生组织细胞的分裂减少且根和胚轴的韧皮部组织严重减少或缺失。 AHK 2 and AHK 3 have also been identified as cytokinin receptors. Control of cell division through cytokinin, appears to be essential for proper cell numbers and phloem formation in the vascular cylinder.
 The SHR gene is expressed in the")
Radial patterning in the ground tissue (A) The SHR gene is expressed in the vascular cylinder (brown), but its activity is required for radial patterning in the adjacent cortex-endodermal initials (orange) and in the endodermis (yellow). SHR protein moves (arrows) to cells surrounding the vascular cylinder, where it induces the expression of SCR and, with the exception of the QC (red), other endodermis specifi c genes. SCR acts downstream of SHR and is required for the periclinal divisions of ground tissue initials which yield the two layers of ground tissue and possibly, for maintaining endodermis identity in the inner layer. B) SHR expression in the embryo. A green fl uorescent protein reporter gene under control of the SHR promoter is expressed exclusively in the central procambium of a triangular stage embryo. Expression remains associated with vascular tissues throughout development. Abbreviations: g, ground tissue; hyp, hypophysis; pc, procambium; pd, protoderm; su, suspensor. (C) SHR: : GFP translational fusion protein is present in the stele, but selectively accumulates in the nuclei of endodermal cells (compare transcriptional fusion in insert). From Nakajima et al. (2001). Abbreviations: Cei, cortex/endodermis initial. Cor, cortex. End, endodermis. Epi, epidermis. Ste, stele. Qc, quiescenter. (D) Ectopic expression of SHR driven by the SCR promoter results extra layers expressing endodermal markers. SHR protein (red) can move one layer beyond its site of production. In normal development (upper panel), it is produced in the vascular tissues (brown) and triggers SCR expression (green) and differentiation events in the overlying ground tissue. SCR is required for periclinal cell divisions subdividing the ground tissue and for proper differentiation of the endodermis (yellow). Expression of SHR in the SCR domain leads to a reiteration of the layer-amplifi cation process and eventually to supernumerary endodermal layers.
- Slides: 64