Per diventare un buon medico occorre essere curiosi

Per diventare un buon medico, occorre essere curiosi

Quale è il meccanismo maggiormente responsabile della produzione di mutazioni spontanee ? ? ?

Errori da replicazione del DNA • il DNA è replicato in 2 -3 ore • ~100, 000 base pairs sono replicate in 1 secondo • è fatale che accadano errori • se non fosse per l'esistenza di un meccanismo biologico di correzione degli errori, le mutazioni prodotte sarebbero assai più numerose, probabilmente di un livello insopportabile (!)

IL DNA E’ UNA MOLECOLA DINAMICA STABILITA' vs INSTABILITA' GENOMA
 divisioni")
ERRORI SPONTANEI CHE AVVENGONO DURANTE LA REPLICAZIONE DEL DNA MUTAZIONI ENDOGENE • 10(17) divisioni cellulari per generare e mantenere una popolazione di 10(14) cellule • per ciascuna divisione cellulare devono essere incorporati 6 x 10(9) nuovi nucleotidi • un individuo "perfetto" richiederebbe una replicazione del DNA priva di errori con una accuratezza maggiore di 6 x 10(26)
 subisca una")
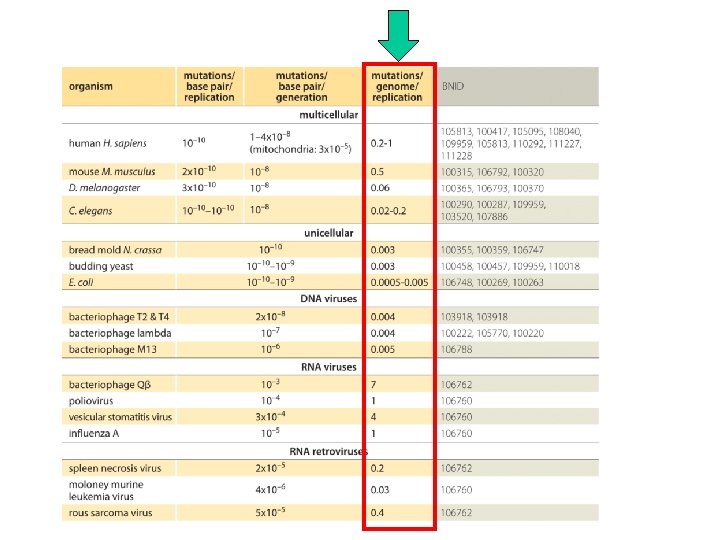
Il tasso di mutazione è equivalente alla probabilità che una sequenza (gene) subisca una mutazione nel corso di una generazione o della produzione di un gamete Geni batterici e virali subiscono mutazioni in media una volta ogni 100 milioni di replicazioni o divisioni cellulari Mais, Drosophila, e uomo dimostrano tassi molto più elevati. Per ogni gamete prodotto si hanno 1 in 1, 000 (106) e 1 in 100, 000 (10 -5) mutazioni/gene (almeno per i geni studiati). Nel topo alcuni geni presentano un tasso di mutazione spontanea ancora più elevato, da 1 in 100, 000 a 1 in 10, 000 (da 10 -5 a 10 -4).

Un esempio di come si misura il tasso delle mutazioni spontanee 11 x 10 -6 mutations per gene (per generation) Può variare tra 10 -8 and 10 -4 in base al gene.

")
Mutation rate In humans, a mutation rate of about 10 -8 mutations/bp/generation (BNID 105813) was inferred from projects where both parents and their children were sequenced at high coverage. Note that the value of the mutation rate is on a per generation basis and is thus the accumulation in the gametes of mutations occurring over several tens of genome replications between fertilization of the egg all the way until the formation of the next generation of gametes. In humans it is estimated that there about 20 -30 genome replications between the fertilized egg and the female gametes (BNID 105585) and about ten times that for males, with large variation depending on age (BNID 105574). With ≈3× 109 bp in the human genome the mutation rate leads to about 10 -8 mutations/bp/generation x 3× 109 bp/genome ≈ 10 -100 mutations per genome per generation (BNID 110293). Using an order of magnitude of 100 replications per generation, we arrive at 0. 1 -1 mutations per genome per replication.

ETEROZIGOSI DA 1: 250 A 1: 1000 BASI SONO DIVERSE TRA DUE INDIVIDUI QUALSIASI I genomi di due individui differiscono per almeno 6 milioni di basi

POLIMORFISMO SEQUENZA POLIMORFICA FREQUENZA > 1% ALLELE

Le mutazioni si generano a caso e quindi colpiscono il genoma a caso ?

Le mutazioni che si generano a caso colpiscono il genoma a caso ? LA FREQUENZA E LA VARIETA’ DI MUTAZIONI NEL DNA CODIFICANTE SONO DIVERSE DAL DNA NON CODIFICANTE

IL TASSO DI TRANSIZIONE NEI GENOMI DI MAMMIFERO E' INASPETTATAMENTE SUPERIORE AL TASSO DI TRANSVERSIONE

C---->T FREQUENZA ELEVATA DERIVANTE DALLA INSTABILITA' PER DEAMINAZIONE SPONTANEA DI C NEL DINUCLEOTIDE Cp. G (TASSO DI MUTAZIONE 8, 5 VOLTE DI ALTRI DINUCLEOTIDI)

Differente grado di conservazione nel tempo di sequenze di tipo diverso LE SEQUENZE CODIFICANTI HANNO IL GRADO PIU ELEVATO DI CONSERVAZIONE

Le mutazioni nella regione codificante Non-conservative VS Conservative mutations DIFFERENTI GRADI DI DEGENERAZIONE CARATTERIZZANO LE DIFFERENTI POSIZIONI NELL'AMBITO DEL CODONE

Differente grado di conservazione nel tempo di sequenze di tipo diverso IL TASSO DI SOSTITUZIONE DI UNA BASE IN SITI NON DEGENERATI E' MOLTO BASSO COERENTEMENTE CON UNA FORTE PRESSIONE SELETTIVA TASSO DI SOSTITUZIONE DI UNA BASE IN SITI DEGENERATI QUATTRO VOLTE E’ MOLTO SIMILE A QUELLO DENTRO INTRONI E PSEUDOGENI PERCHE’ SELETTIVAMENTE NEUTRE TASSO DI SOSTITUZIONE DI UNA BASE IN SITI DEGENERATI DUE VOLTE E’ INTERMEDIO PERCHÉ ALCUNE SOSTITUZIONI SONO CONSERVATIVE

Mutazioni puntiformi nel codone di Tyr Thr Tyr Ala protein 5’-ACG UAU GCG -3’ 1 st nucleotide changes 3 rd nucleotide changes GAU = Asp UGU = Cys AAU = Asn UCU = Ser CAU = His UUU = Phe Missense m. RNA UAG = stop UAA = stop UAC = Tyr Missense Non-conservative VS Conservative mutations nonsense silent


SITI NON DEGENERATI=TUTTE E 3 LE POSSIBILI SOSTITUZIONI SONO NON SINONIME • PRIMA BASE 53/61 • SECONDA BASE 61/61 • TERZA BASE 2/61 • TOTALE 65%

SITI DEGENERATI DUE VOLTE = UNA DELLE TRE POSSIBILI SOSTITUZIONI E’ UN SINONIMO PRIMA BASE 8/61 TERZA BASE 27/61 TOTALE 19%

SITI DEGENERATI QUATTRO VOLTE = TUTTE E 3 LE POSSIBILI SOSTITUZIONI SONO SINONIME • TERZA BASE 32/61 • TOTALE 16%

SOSTITUZIONI CONSERVATIVE E NONCONSERVATIVE

SOSTITUZIONI CONSERVATIVE E NONCONSERVATIVE LA STRUTTURA DEL CODICE GENETICO E IL GRADO CON CUI UN AMINOACIDO E’ FUNZIONALMENTE SIMILE AD UN ALTRO INFLUISCONO SULLA RELATIVA MUTABILITÀ DEI SINGOLI AMINOACIDI (CERTI AMINOACIDI NON SONO FACILMENTE SOSTITUIBILI AD ES. CISTEINA AL CONTRARIO SERINA E TREONINA SONO TRA I PIU' MUTABILI)
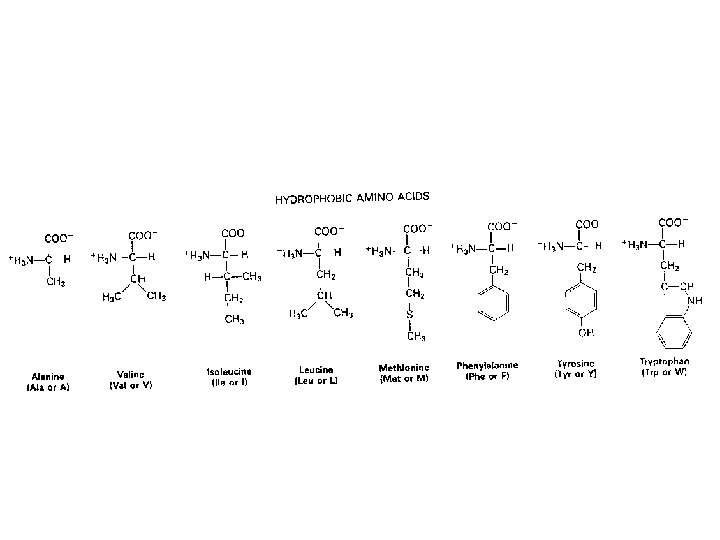
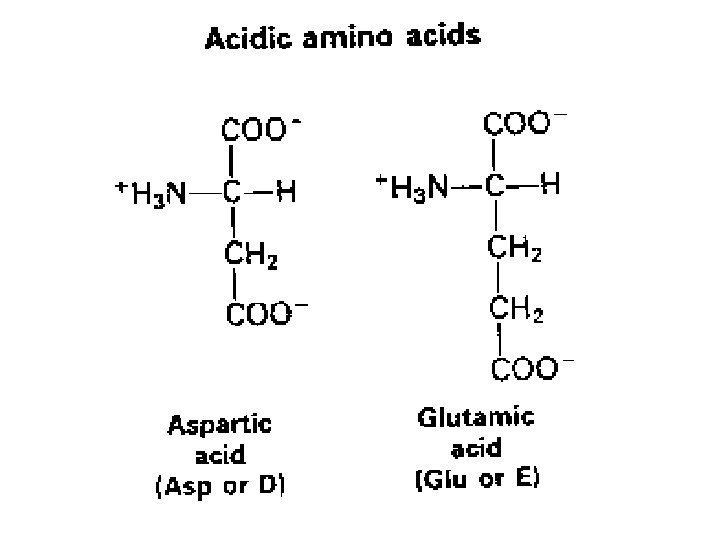
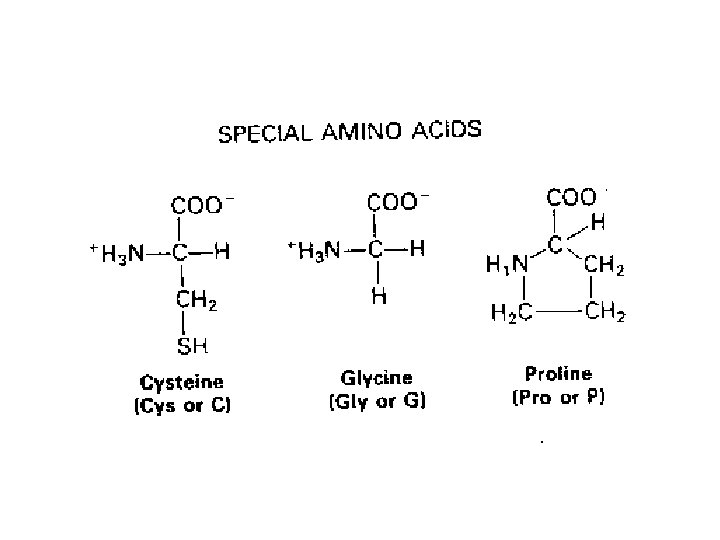

Il tasso e il tipo di sostituzioni variano tra geni differenti I geni che codificano proteine mostrano enormi variazioni nel tasso di sostituzioni non sinonime FATTORI CHE DETERMINANO DIFFERENZE • Funzione biologica delle proteine (sequenze estremamente conservate) • Specie animale (roditore ----> scimmia -----> uomo) • Sesso

I tassi di mutazione sono più elevati nei maschi per il maggior numero di divisioni delle cellule germinali

Femmina: durante il V mese di sviluppo la popolazione totale di cellule germinali raggiunge la quota massima di 6, 8 x 106 cellule. Sono necessari 22 cicli replicativi. Per diventare cellula uovo aggiungi altre due divisioni (meiotiche). Totale 24 cicli.

Maschio: fino alla pubertà di anni 13 si producono 1, 2 x 109 cellule germinali (spermatogoni) mediante 30 cicli replicativi. Dopo la pubertà gli spermatogoni affrontano ogni anno un totale di 23 cicli. Lo spermatogonio matura a spermatozoo mediante 5 cicli. Totale 30 + 23 n + 5 (n=età in anni dopo la pubertà).

IL TASSO COMPLESSIVO DI MUTAZIONE NEL DNA CODIFICANTE E' MOLTO INFERIORE RISPETTO AL DNA NON CODIFICANTE LA PRESSIONE SELETTIVA RIDUCE NON SOLO LA FREQUENZA COMPLESSIVA MA ANCHE LO SPETTRO DELLE POSSIBILI MUTAZIONI NEL DNA CODIFICANTE MUTAZIONI SILENTI, SENSE, E NON-SENSE HANNO DIVERSO EFFETTO E SI DISTRIBUISCONO CON DIFFERENTI FREQUENZE

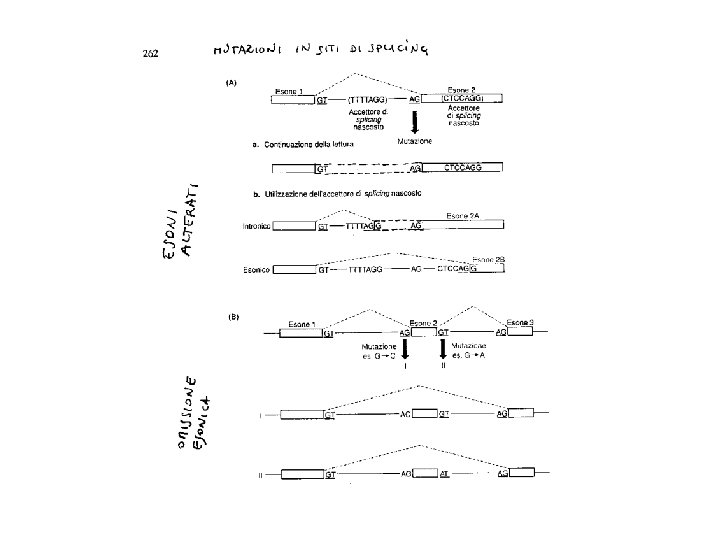
LE MUTAZIONI CHE INTERESSANO LO SPLICING alterano una sequenza conservata necessaria per lo splicing normale, ma alcune si verificano in sequenze normalmente non richieste per lo splicing

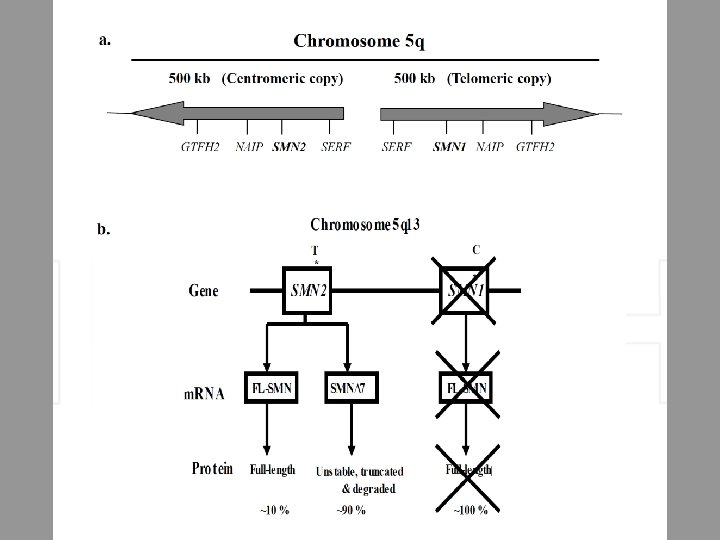
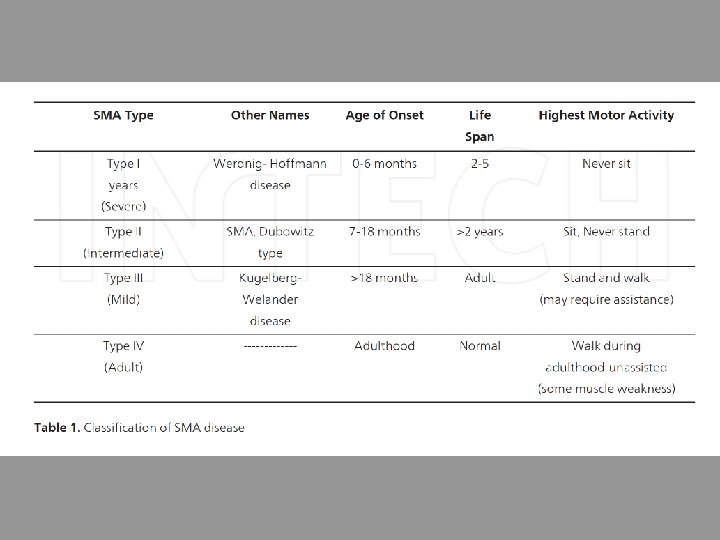
Il caso del gene SMN • Spinal-Muscolar-Atrophy-determining gene Survival of Motor Neurons (SMN, 3. 5 KB) • Part of 500 Kb inverted duplication 5 q 13 • SMA patients homozygous deletion SMN 1 gene • Deletion of SMN 2 is not associated with SMA (Melki et al. , 1994; Lefebvre et al. , 1995; Burglen et al. , 1996)

Genetics of the disease The SMA disease causing SMN 1 gene maps to a complex genomic region of chromosome 5 q 13. 1. This region is characterized by an inverted duplication of the element with 4 genes (SMN, neuronal apoptosis inhibitor protein {NAIP}, SERF and GTFH 2) present in telomeric and centromeric copies. In 1995, it was reported that homozygous deletions of the SMN 1 gene were likely the cause of 95% of SMA patients. All SMA patients have one or more copies of a nearly identical gene, SMN 2. These two genes are distinguished by five nucleotide changes in exon 7 and 8. The critical nucleotide difference which makes SMN 2 only partially functional is a C to T transition at position 6 of exon 7. This change leads to the exclusion of exon 7 in the majority of transcripts. This m. RNA is subsequently translated to form an unstable truncated non-oligomerizing isoform of SMN protein. However, SMN 2 gene still produces 5 -10% functional full length SMN transcripts. The SMN 2 gene is present in variable copy numbers in the population; all SMA patients have one or more copy of the SMN 2 gene which, due to its partial functionality, acts as a positive disease modifier. There is thus an inverse correlation between the number of SMN 2 gene (which can produce between 10 -50% of SMN protein depending on copy number) and the severity of the disease. Low levels of SMN protein allows embryonic development but is not enough, in the long term, to allow motor neurons to survive in the spinal cord. Type I patients usually have 2 copies whereas Type II have 3 copies of SMN 2. Type III and IV have 3 -4 copies of the SMN 2 gene. Individuals with 5 or more copies of the SMN 2 gene, despite having no functional SMN 1 gene are completely asymptomatic and are protected against the disease manifestation.

Normal Individual SMN 2 SMN 1 SMA Patients SMN 2 SMN 1
")
Quando una mutazione silente non è silente Distrofia muscolare del cingolo (gene calpaina 3)

Mutazioni che introducono un codone di terminazione prematura • Mutazioni non sense • Inserimenti e delezioni (un codone di STOP entro circa 100 nt) • Omissione di esoni • ----> m. RNA instabile (++ frequente) • ----> Polipeptide tronco (instabile)

Mutazioni che coinvolgono sequenze ripetute Sequenze ripetute di nucleotidi sono presenti in elevato numero di copie nel DNA, hanno strutture diverse (minisatelliti, microsatelliti, VNTR, etc. ), e sono la causa più frequente di inserzioni, delezioni e fusioni
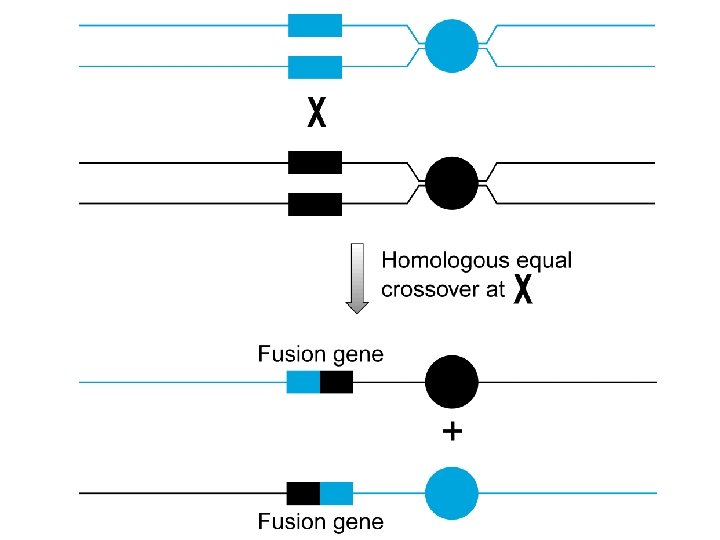
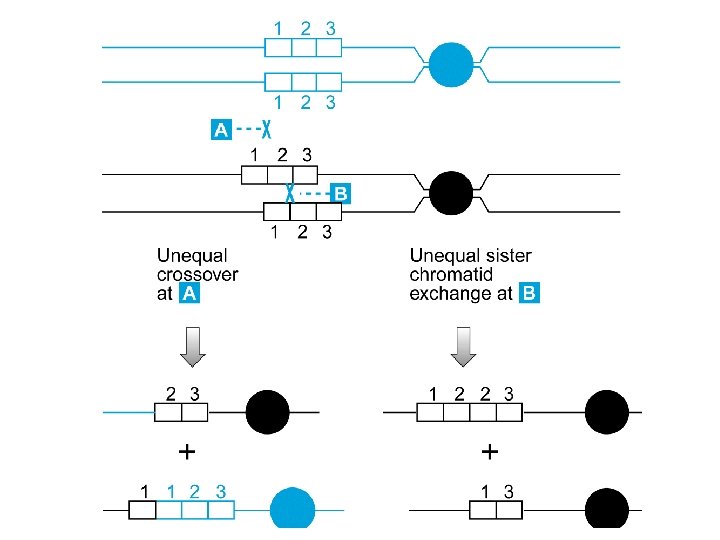

Duplicazione genica
")
Sindrome di Kearns-Sayre (miopatia mitocondriale)


Background • Rare neuromuscular disorder • • Effects mitochondria • • • Large duplications or deletions in mitochondrial DNA (>1000 bp) results in under or over production of mitochondrial gene products 1/3 cases 4977 bp deletion Randomly effects people • • Only 226 cases reported in literature only isolated reports of any kind of inheritance pattern Prognosis • • Depends on severity Severe cases 85% muscle mitochondrial genome affected

Symptoms • Onset before 20 and includes • • • Immobile or limited eye movement Abnormal pigment in eyes Cardiac problems or congestive heart failure Muscle and skeletal weakness Short stature Hearing Ataxia (coordination problems) Diabetes Dementia and other mental illnesses Affects systems with higher energy requirements most
 in Kearns-Sayre")
Fibre rosse sfilacciate (Ragged red fibers) in Kearns-Sayre

Conversione genica Trasferimento non reciproco di sequenze informative tra una coppia di sequenze non alleliche (conversione genica interlocus) o alleliche (c. g. interallelica) Relativamente frequenti a livello di DNA ripetuto in tandem
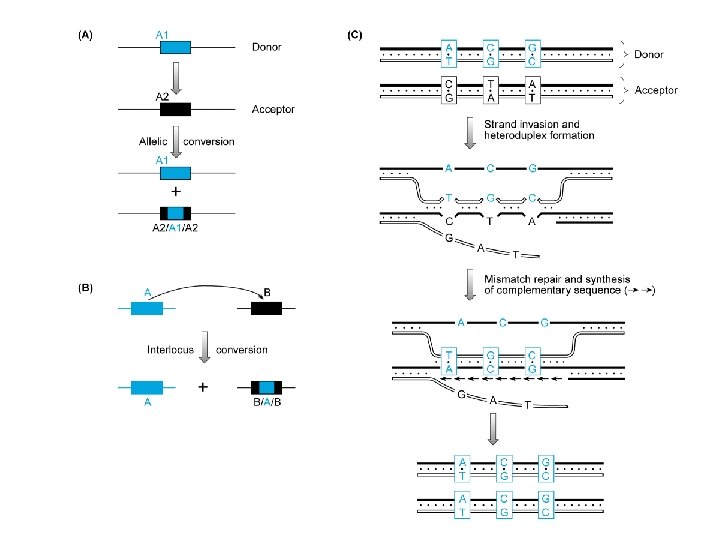

Steroido-21 -idrossilasi Scambi gene-pseudogene Congenital adrenal hyperplasia autosomal recessive

prima dopo

Inversioni prodotte dalla ricombinazione intracromatidica tra ripetizioni invertite

Emofilia A: un problema di inversione • Il gene del fattore VIII della coagulazione si trova sul cromosoma X ed è la causa più importante di emofilia • F 8 è un gene molto lungo: all'interno dell'introne 22 sono presenti un gene più piccolo, F 8 A insieme a una seconda copia collocata alcune centinaia di migliaia di cb, sulla catena opposta • Quindi i due geni che sono assai simili tra loro hanno orientamento opposto. • Accade con una certa frequenza che nel corso della meiosi i due geni vanno incontro a ricombinazione intra-allelica per appaiamento errato • Questa mutazione da ragione del 45% dei casi di emofilia A • Quasi tutti i nuovi casi insorgono durante la meiosi nel maschio: infatti nella femmina i due cromosomi omologhi sono appaiati durante la meiosi e questo fatto pare inibire la inversione

Emofilia A

Sequenze molto brevi di DNA ripetuto sono spesso coinvolte nella produzione di mutazioni importanti

fibrosi cistica poliposi adenomatosa del colon xeroderma pigmentoso beta-talassemia

- Slides: 66