PARED CELULAR INTRODUCCIN La mayor parte de los











 y la Llys. Ambos son diaminoácidos. El m-DAP se")

 Puente pentaglicina (algunas Gram positivas")





,")






 Pared especial de ciertas Gram-positivas: Nocardia, Mycobacterium")
, 4")
- Slides: 34
PARED CELULAR INTRODUCCIÓN La mayor parte de los procariotas posee una pared celular (P. C. ) rígida rodeando al protoplasto. Las excepciones son los micoplasmas (dentro del dominio Bacteria) y algunas arqueas, como Thermoplasma. Al microscopio electrónico se puede observar como una capa en íntimo contacto con la membrana citoplásmica, con un espesor que oscila entre 10 y 80 nm (según especies) -frente a los 8 nm de la membrana celular- , y con una estructura más o menos compleja, según los tipos bacterianos. a) Las paredes celulares más frecuentes en eubacterias siguen dos modelos alternativos que, como veremos comparten un componente común: paredes de tipo Gram-positivo o de tipo Gram-negativo. b) Unas pocas eubacterias (como las del gén. Planctomyces) poseen paredes a base de proteínas. c) Las Arqueas poseen paredes diferentes a las de eubacterias y se pueden agrupar en diversos tipos. La pared celular bacteriana está hecha de peptidoglicano (también denominado mureína), que está formado por cadenas de polisacárido entrecruzadas por péptidos inusuales que contienen aminoácidos. D
. Las paredes celulares bacterianas son diferentes de las paredes de plantas y hongos que están hechas de celulosa y quitina, respectivamente. También son diferentes de las paredes de Archaea, que no contienen peptidoglicano. La pared celular es esencial para la supervivencia de muchas bacterias En las bacterias Gram-positivas la pared celular contiene una capa gruesa de peptidoglicano además de ácidos teicoicos, que son polímeros de glicerol o ribitol fosfato. Los ácidos teicoicos se unen al peptidoglocano o a la membrana citoplasmática. En las bacterias Gram-negativas la capa de peptidoglicano es relativamente fina y se encuentra rodeada por a una segunda membrana lípida exterior que contiene lipopolisacáridos y lipoproteínas. La capa de petidoglicano se une a la membrana externa por medio de lipoproteínas. La mayoría de las bacterias tienen una pared celular Gram-negativa y solamente Firmicutes y Actinobacteria (conocidas previamente como bacterias Gram-positivas de contenido GC bajo y bacterias Grampositivas de contenido GC alto, respectivamente) tienen paredes Grampositivas
• La tinción de Gram es la más importante entre las tinciones llamadas diferenciales (aquellas que no tiñen de la misma manera a todos los tipos de bacterias), esta tinción consta de varios pasos: • 1. Imaginemos un frotis en porta de vidrio con una muestra de varios tipos de bacterias, que se han fijado por calor. En la primera fase, esta preparación de trata con un primer colorante llamado violeta cristal o violeta de genciana. • 2. A continuación se añade una solución de lugol (yodo-ioduro), que actúa como “mordiente”, formando una laca relativamente resistente con el violeta. En este momento todas las bacterias están teñidas de color violeta • 3. Ahora realizamos una descoloración diferencial con etanol o una mezcla de etanol y acetona. Lo que ocurre es que algunas bacterias (las que llamaremos Gram-negativas) pierden el color violeta (quedarían práctimante transparentes al microscopio), mientras que otras (las Grampositivas) resisten el tratamiento, y retienen el colorante violeta. • 4. Finalmente, tratamos el porta con un segundo colorante (colorante de contraste): se trata de un colorante de color rojo o rosa, como la fucsina o la safranina. Este colorante tiñe ahora a las bacterias previamente descoloradas por el etanol. Por lo tanto, el resultado en la observación al microscopio es que las bacterias Gram-positivas se ven de color violeta, y las Gram-negativas de color rosa o rojo.
Tinción de Gram
3. Ahora realizamos una descoloración diferencial con etanol o una mezcla de etanol y acetona. Lo que ocurre es que algunas bacterias (las que llamaremos Gram-negativas) pierden el color violeta (quedarían prácticamente transparentes al microscopio), mientras que otras (las Gram-positivas) resisten el tratamiento, y retienen el colorante violeta. 4. Finalmente, tratamos el porta con un segundo colorante (colorante de contraste), de color rojo o rosa, como la fucsina o la safranina. Este colorante tiñe ahora a las bacterias previamente descoloradas por el etanol. Por lo tanto, el resultado en la observación al microscopio es que las bacterias Gram-positivas se ven de color violeta, y las Gram-negativas de color rosa o rojo. Como veremos enseguida, esta diferencia de comportamiento ante la tinción de Gram refleja el hecho de que ambos tipos de eubacterias poseen dos tipos estructuralmente diferentes de pared celular, aunque ambas posean en común la posesión de peptidoglucano. A continuación estudiaremos con cierto detalle la pared celular de eubacterias.
BACTERIAS GRAM POSITIVAS
BACTERIAS GRAM NEGATIVAS
BACTERIAS ÁCIDO-ALCOHOL RESISTENTES
PAREDES DE LAS EUBACTERIAS
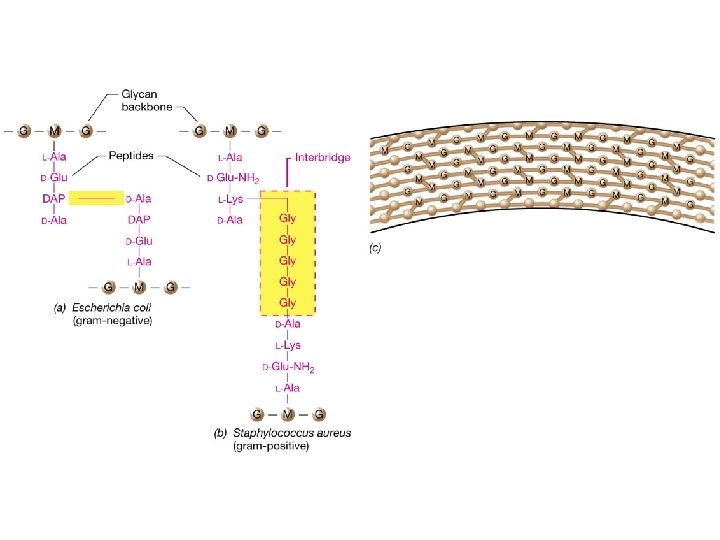
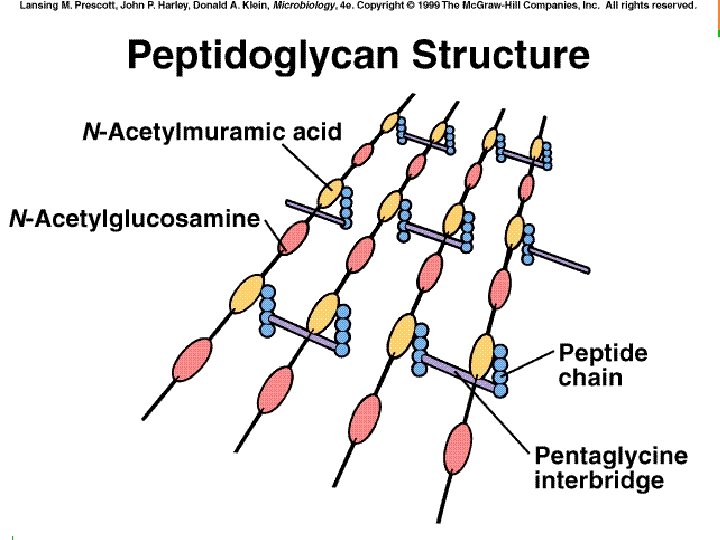
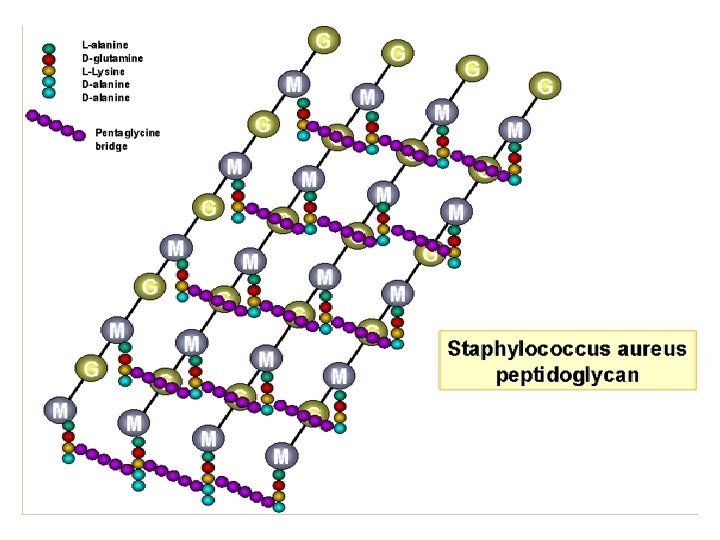
COMPOSICIÓN QUÍMICA Y ESTRUCTURA BÁSICAS DEL PEPTIDOGLUCANO Está formado por repeticiones de una unidad disacarídica fundamental unida a su vez a un tetrapéptido. Distintas cadenas (formadas por el esqueleto de azúcares) se unen entre sí por determinados enlaces peptídicos entre tetrapétidos de cadenas diferentes. La unidad disacarídica repetitiva: consiste en Nacetilglucosamina (NAG) unida por enlace ß(1 4) a N-acetilmurámico (NAM). Obsérvese que el NAM es el 3 -O-D-lactil-éter de la NAG (o sea, se deriva de unir el ácido D-láctico con el OH del C-3 de la NAG). Las distintas unidades disacarídicas se van uniendo entre sí por enlaces ß(1 4) entre el NAM de una unidad y la NAG de la siguiente. Este enlace es susceptible a la rotura catalizada por el enzima lisozima. El número de repeticiones (n) puede oscilar entre 10 y 100. La cadena tetrapeptídica: Desde el grupo carboxilo de cada ácido NAM, y mediante un enlace amido, se encuentra unido el tetrapéptido. Un tetrapéptido típico de muchas bacterias es: L-alanina - D-glutámico - meso-diaminopimélico - D-alanina
Fórmula de un peptidoglucano típico. Observar: • La unidad disacarídica NAG-NAM • Las distintas unidades disacarídicas se unen entre sí por enlaces β(1à 4) entre el NAM de una unidad y el NAG de la siguiente. Este enlace es susceptible de rotura por lisozima en muchas bacterias • El tetrapéptido, con su alternancia de aminoácidos en L y en D
Comparación entre el meso-diaminopimélico (m-DAP) y la Llys. Ambos son diaminoácidos. El m-DAP se puede considerar como una Lys en la que el –H terminal de la cadena lateral está sustituido por un carboxilo (-COOH) m-DAP L-lys
LA ESTRUCTURA GLOBAL: Las distintas cadenas polisacarídicas, con sus respectivos tetrapéptidos, se unen entre sí por medio de puentes o enlaces peptídicos, entre un aminoácido de una cadena (p. ej. , el aminoácido nº 3, como el meso-DAP del ejemplo) y otro aminoácido de una cadena adyacente (la D-ala terminal). De este modo, la estructura global es una sola macromolécula gigante que envuelve al protoplasto, formando un sáculo rígido, a modo de tejido continuo, que tiene el volumen y la forma de la bacteria respectiva
Ejemplos de entrecruzamientos en el peptidoglucano Directo (muchas Gram-negativas) Puente pentaglicina (algunas Gram positivas
Comparación de las envolturas celulares bacterianas. Arriba: Bacteria Grampositiva. 1 -membrana citoplasmática, 2 peptidoglicano, 3 fosfolípidos, 4 proteínas, 5 -ácido lipoteicoico. Abajo: Bacteria Gramnegativa. 1 -membrana citoplasmática (membrana interna), 2 espacio periplasmático, 3 -membrana exterior, 4 fosfolípidos, 5 peptidoglicano, 6 lipoproteína, 7 proteínas, 8 lipopolisacáridos, 9 porinas.
BACTERIA GRAM NEGATIVA Las bacterias Gram-negativas presentan dos membranas lipídicas entre las que se localiza una fina pared celular de peptidoglicano, mientras que las bacterias Gram-positivas presentan sólo una membrana lipídica y la pared de peptiglicano es mucho más gruesa. Al ser la pared fina, no retiene el colorante durante la tinción de Gram. Muchas especies de bacterias Gram-negativas causan enfermedades. Los cocos Gram-negativos causan la gonorrea (Neisseria gonorrhoeae), meningitis (Neisseria meningitidis) y síntomas respiratorios (Moraxella catarrhalis), entre otros. Los bacilos Gram-negativos incluyen un gran número de especies. Algunos de ellos causan principalmente enfermedades respiratorias (Hemophilus influenzae, Klebsiella pneumoniae , Legionella pneumophila, Pseudomonas aeruginosa), enfermedades urinarias (Escherichia coli, Proteus mirabilis, Enterobacter cloacae, Serrati marcescens) y enfermedades gastrointestinales (Helicobacter pylori, Salmonella enteritidis, Salmonella typhi). Otros están asociadas a infecciones nosocomiales (Acinetobacter baumanii
EL PEPTIDOGLUCANO DE BACTERIAS GRAM-NEGATIVAS En la mayor parte de Gram-negativas el peptidoglucano corresponde a la composición y estructura que acabamos de describir. Sin embargo, en las espiroquetas, el diaminoácido en posición 3, en vez de ser meso-DAP, está sustituido por la L-ornitina (que también es un diaminoácido). El enlace entre cadenas polisacarídicas se realiza normalmente mediante unión peptídica directa entre el grupo carboxilo de la D-ala terminal y el grupo amino del meso-DAP. Ahora bien, en este enlace participan solamente el 50% de los tetrapéptidos. Los demás péptidos no participan en enlaces, y entre estos últimos se encuentran incluso dipéptidos y tripéptidos. El resultado es una capa simple de PG (de 1 nm de espesor), a modo de malla floja, y con grandes poros (los “huecos” dejados por las zonas donde no hay enlace peptídicos). Ello explica el comportamiento de las bacterias Gram-negativas en la tinción de Gram: al añadir el alcohol, se produce una deshidratación que tiende a contraer la estructura del PG, pero los poros son grandes y por ellos sale el primer colorante (el violeta de genciana). El ulterior tratamiento de la preparación con el colorante de contraste (fuchsina o safranina) tiñe a estas bacterias de rojo.
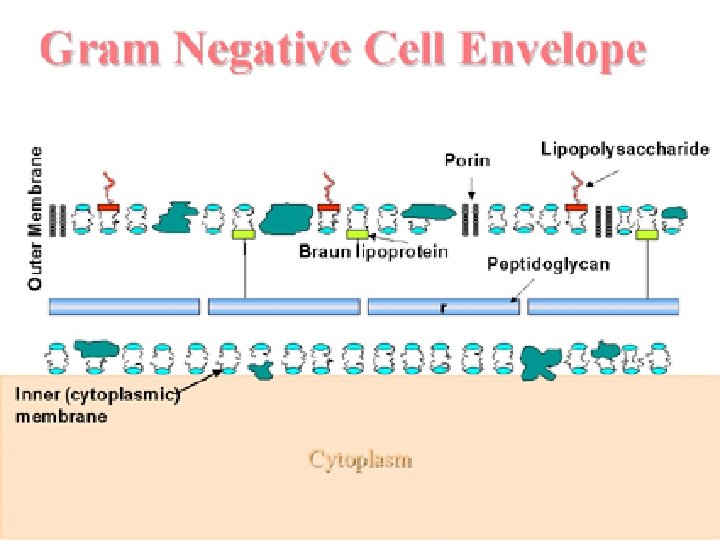
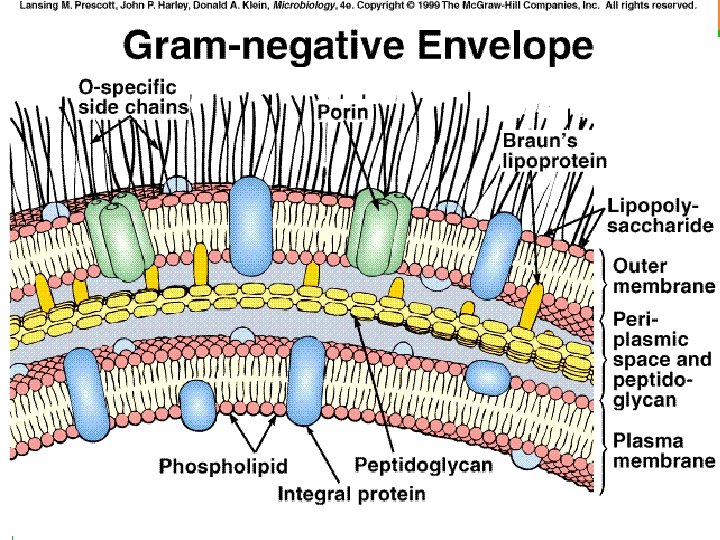
La envoltura celular de las bacterias Gram-negativas está compuesta por una membrana citoplasmática (membrana interna), una pared celular delgada de peptidoglicano, que rodea a la anterior, y una membrana externa que recubre la pared celular de estas bacterias. Entre la membrana citoplasmática interna y la membrana externa se localiza el espacio lásmico relleno de una sustancia denominada periplasma, la cual contiene enzimas importantes para la nutrición en estas bacterias. La membrana externa contiene diversas proteínas, siendo una de ellas porinas o canales proteícos que permiten el paso de ciertas sustancias. También presenta unas estructuras llamadas lipopolisacáridos (LPS), formadas por tres regiones: el polisacárido O (antígeno O), una estructura polisacárida central (KDO) y el lípido A (endotoxina). Las bacterias Gram-negativas pueden presentar una capa S que se apoya directamente sobre la membrana externa, en lugar de sobre la pared de peptidoglicano como sucede en las Gram-positivas. Si presenten flagelos, estos tienen cuatro anillos de apoyo en lugar de los de las bacterias Grmapositivas porque tienen dos membranas. No presentan ácidos teicoicos ni ácidos lipoteicoicos, típicos de las bacterias Gram-positivas. Las lipoproteínas se unen al núcleo de polisacáridos, mientras que en las bacterias Grampositivas estos no presentan lipoproteínas. La mayoría no forma endosporas (Coxiella burnetti, que produce estructuras similares a las endosporas, es una notable excepción).
PARED GRAM NEGATIVA Otra imagen general de las envueltas de una bacteria Gramnegativa (vale la pena recordar. . . !. Observa la estructura general de la pared celular: • el espacio periplásmico contiene el PG, el cual se une covalentemente con la lipoproteína de Braun, que a su vez contacta con la lámina interna de la membrana externa • la membrana externa es una bicapa proteolipídica muy asimétrica: • su lámina externa contiene el LPS (observa las cadenas laterales proyectadas hacia el exterior) • atravesando la membrana externa hay varios tipos de proteínas, incluyendo los trímeros de porinas, con sus correspondientes canales internos, que solo dejan pasar moléculas hidrófilas por debajo de cierto tamaño
El lipopolisacárido El lípido A: Dos unidades de glucosamina unidas por enlace ß(1à 6), pero donde todos los grupos -OH (menos uno) y -NH 2 están sustituidos (unidos a otras moléculas): Obsérvese que A. G. saturados (C-14): betahidroximirístico ·existen 5 (a veces 6) ácidos grasos, todos ellos saturados, con predominio de ß-hidroximirístico (un ácido graso C 14). · El -OH original en 4´ está sustituido por arabinosaminafosfato. · El -OH en 1 está sustituido por fosforil-etanolamina (a veces pirofosforil-etanolamina). El oligosacárido medular (también llamado corazón o núcleo): se une al lípido A a través del -OH en 3´. Se pueden considerar dos fracciones: · núcleo interno, a base de dos tipos de azúcares exclusivos de Gram-negativas: 2 -ceto-3 -desoxioctónico (KDO) y L-glicero-D-manoheptosa (Hep). . Alguna de las Hep y alguno de los KDO pueden a su vez estar unidos a fosforil-etanolamina (o pirofosforil-etanolamina). Esta región es muy rica en grupos cargados, especialmente con carga negativa (de los fosfatos y KDO). · La fracción del núcleo externo está constituida a base de hexosas (glucosa, galactosa, NAG, y a veces algunas hexosas más raras). Unidad repetitiva de la cadena lateral Cadena lateral específica: polisacárido repetitivo, que se proyecta hacia el exterior celular. Consiste en la repetición (n<40 veces) de unidades tri-, tetra- o pentasacarídicas (en estos dos últimos casos uno de los azúcares de cada repetición queda lateral respecto del esqueleto lineal que forman los demás). Núcleo externo Núcleo interno Glucosaminaß(1 6)glucosamina, con –OH en 1 sustituido con –Petanolamina
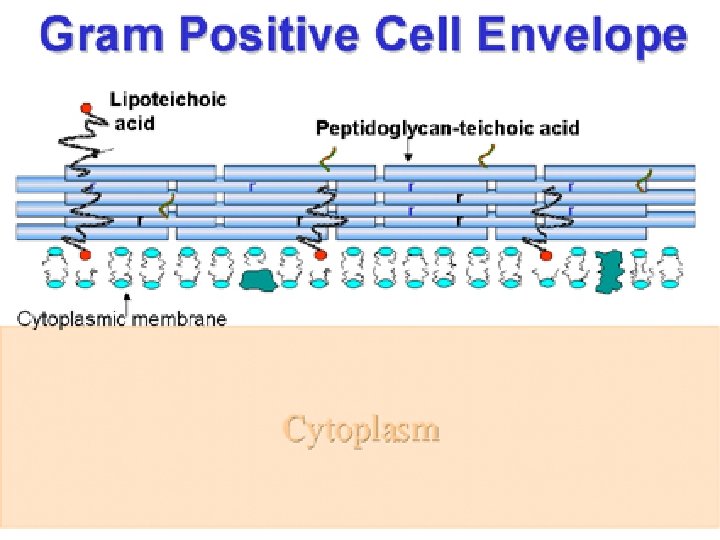
Bacteria Gram positiva La envoltura celular de las bacterias Gram-positivas comprende la membrana citoplasmática y una pared celular compuesta por una gruesa capa de peptidoglicano, que rodea a la anterior. La pared celular se une a la membrana citoplasmática mediante moléculas de ácido lipoteicoico. La capa de peptidoglicano confiere una gran resistencia a estas bacterias y es la responsable de retener el tinte durante la tinción de Gram. A diferencia de las Gram-negativas, estas bacterias no presentan una segunda membrana lipídica externa. Incluyen especies tanto móviles como inmóviles con forma de bacilo (Bacillus, Clostridium, Corynebacterium, Lactobacillus, Listeria) o coco (Staphylococcus, Streptococcus); con gruesas paredes celulares o sin ellas (Mycoplasma). Algunas especies son fotosintéticas, pero la mayoría son heterótrofas. Muchas de estas bacterias forman endosporas en condiciones desfavorables.
La célula bacteriana está rodeada por una envoltura que observada al microscopio electrónico, se presenta como una capa gruesa y homogénea denominada pared celular. Entre la membrana plasmática y la pared celular se encuentra el periplasma o espacio periplasmático, luego en sección (corte) se observa una estructura semejante a dos líneas paralelas formada separando una capa menos densa; esto corresponde a la llamada envoltura bacteriana. En el interior de la membrana plasmática se encuentra el citoplasma que está constituido por una disolución acuosa, el citosol, en el cual se encuentran ribosomas y otros agregados de macromoléculas, y en el centro se ubica la zona menos densa llamada nucleoide, que contiene una madeja de hebras difícil de resolver (distinguir) y cuyo principal componente es el ADN. La pared externa de la envoltura celular de una bacteria Gram positiva tiene como base química fundamental el peptidoglucano, que es un polímero de Nacetil-2 -D-glucosamina, unido en orientación ß-1, 4 con N-acetil murámico, a éste se agregan por el grupo lactilo 4 ó más aminoácidos.
Esta molécula se polimeriza gran cantidad de veces, de modo que se forma una malla especial, llamada sáculo de mureína. Dicho compuesto es de vital importancia para conservar la forma y darle rigidez a la célula bacteriana (si este compuesto no existiese, la célula reventaría debido a su gran potencial osmótico). Las siguientes características son presentes generalmente en una bacteria Grampositiva: Membrana citoplasmática. Capa gruesa de peptidoglicano. ácido teicoicos y lipoteicoicos, sirven como agentes quelantes y en ciertos tipos de adherencia. Polisacáridos de cápsula Si un flagelo está presente, este contiene dos anillos como soporte en oposición a los cuatro que existen en bacterias Gram-negativas porque las bacterias Grampositivas tienen solamente una capa membranal. Tanto las bacterias Gram-positivas y las Gram-negativas pueden tener una membrana llamada capa S. En las bacterias Gram-negativas, la capa S está unida directamente a la membrana externa. En bacterias Gram-positivas, la capa S está unida a la capa de péptidoglicano. Es único a las bacterias Gram-positivas la presencia de ácidos teicoicos en la pared celular. Algunos ácidos teicoicos particulares, los ácidos lipoteicoicos, tienen un componente lipídico y pueden asistir en el anclaje del péptidoglicano, en tanto el componente lipídico sea integrado en la membrana.
Un ácido ribitol-teicoico
Ácidos teicurónicos: Ciertas bacterias Gram-positivas, cuando se someten a un régimen de limitación de fosfato son incapaces de sintetizar ácidos teicoicos, pero en su lugar producen ácidos teicurónicos. Los teicurónicos consisten en polímeros aniónicos formados por la alternancia de ácidos urónicos (que tienen grupos -COOH libres) y aminozúcares como la N-acetil-galactosamina. Ácidos lipoteicoicos: Están presentes en todas las bacterias Gram-positivas, aun en condiciones de carencia de fosfato. Se trata simplemente de ácidos glicerol-teicoicos que se encuentran unidos a la membrana citoplásmica, concretamente se unen por enlace fosfodiéster con glucolípidos de membrana, mientras que el otro extremo de la cadena queda expuesto al exterior.
Otra imagen que muestra la estructura global de la pared de Gram-positivas. Observar que el PG está inmerso en la matriz a base de: • Teicoicos (unidos al PG) • Lipoteicoicos (unidos a glucoproteínas de la pared celular) Ambos tipos de polímeros pueden sobresalir de la pared hacia fuera. Funciones de los polímeros de la matriz: · Parece ser que su papel principal es suministrar una carga neta negativa a la pared celular, lo que permite captar cationes divalentes (p. ej. , Mg++), que a su vez se necesitan para muchas actividades enzimáticas de la membrana citoplásmica o del espacio periplásmico, que participan de la morfogénesis y división de la pared celular. ·Como ya dijimos, los ácidos teicoicos y teicurónicos son buenos antígenos. Cuando no están cubiertos por estructuras más externas (como cápsulas), constituyen el antígeno somático O de las bacterias Gram-positivas. ·Finalmente, en algunas bacterias, sumistran, junto con el PG, receptores específicos para la adsorción de ciertos bacteriófagos.
PARED DE LAS BACTERIAS ÁCIDO-ALCOHOL RESISTENTES (AAR) Pared especial de ciertas Gram-positivas: Nocardia, Mycobacterium Resisten la decoloración con clorhídrico-etanol ( ácido-alcohol resistentes) Esta propiedad deriva de: Ácidos micólicos, Glucolípidos, Y Ceras Ácidos micólicos Son ß-hidroxiácidos grasos ramificados en α, de cadena muy larga Forman parte de un esqueleto muy peculiar de la pared celular: PG arabinogalactano ácidos micólicos Otros lípidos de en la pared de bacterias AAR Glucolípidos: Micolatos de trehalosa Sulfolípidos de trehalosa Micósidos (unión éster entre micólicos y azúcares) Ceras: unión de micólicos con alcoholes ramificados de cadena larga (C 30 -C 34) Papeles conferidos por la pared AAR Aspecto y consistencia cérea de las colonias en placas de Petri En líquidos crecen formando grumos Gran impermeabilidad Resistencia a desecación Resistencia a agentes antibacterianos Detergentes Oxidantes Ácidos y bases
PARED CELULAR ACIDO-ALCOHOL RESISTENTE: 1 -lípidos externos, 2ácido micólico, 3 -polisacáridos (arabinogalactano), 4 -peptidoglicano, 5 -membrana plasmática, 6 -lipoarabinomanano (LAM), 7 -fosfatidilinositol manosido, 8 -esquema de la pared celular.